
วิวัฒนาการกระดูกหูสำหรับได้ยินของสัตว์เลี้ยงลูกด้วยนม (อังกฤษ: evolution of mammalian auditory ossicles) เป็นเหตุการณ์ทางวิวัฒนาการที่มีหลักฐานยืนยันดีที่สุด และสำคัญที่สุด เหตุการณ์หนึ่ง โดยมีทั้งซากดึกดำบรรพ์ช่วงเปลี่ยนสภาพ (transitional fossil) จำนวนมากและตัวอย่างที่เยี่ยมของกระบวนการ exaptation คือการเปลี่ยนจุดประสงค์ของโครงสร้างที่มีอยู่แล้วในระหว่างวิวัฒนาการ
ในสัตว์เลื้อยคลาน แก้วหูจะเชื่อมกับหูชั้นในผ่านกระดูกท่อนเดียว คือ columella ในขณะที่ขากรรไกรล่างและบนจะมีกระดูกหลายท่อนที่ไม่พบในสัตว์เลี้ยงลูกด้วยนม คือ ในช่วงวิวัฒนาการของสัตว์เลี้ยงลูกด้วยนม กระดูกชิ้นหนึ่งของขากรรไกรล่างและบน (articular และ quadrate) หมดประโยชน์โดยเป็นข้อต่อ และเกิดนำไปใช้ใหม่ในหูชั้นกลาง ไปเป็นตัวเชื่อมกับกระดูกโกลนที่มีอยู่แล้ว รวมกันกลายเป็นโซ่กระดูกสามท่อน (โดยเรียกรวมกันว่ากระดูกหู [ossicles]) ซึ่งถ่ายทอดเสียงได้อย่างมีประสิทธิภาพกว่า และดังนั้นช่วยให้ได้ยินได้ดีกว่า ในสัตว์เลี้ยงลูกด้วยนม กระดูกหูสามท่อนนี้เรียกว่า กระดูกค้อน กระดูกทั่ง และกระดูกโกลน ตามลำดับ สัตว์เลี้ยงลูกด้วยนมและสัตว์ปีกยังต่างจากสัตว์มีกระดูกสันหลังอื่น ๆ เพราะมีคอเคลียที่วิวัฒนาการเกิดขึ้น
หลักฐานว่า กระดูกค้อนและกระดูกทั่งมีกำเนิดเดียวกัน (homologous) กับกระดูก articular และ quadrate ของสัตว์เลื้อยคลานเบื้องต้นมาจากคัพภวิทยา แล้วต่อมา การค้นพบซากดึกดำบรรพ์ช่วงเปลี่ยนสภาพมากมายก็ได้ยืนยันข้อสรุปนี้ โดยให้ประวัติการเปลี่ยนสภาพอย่างละเอียด ส่วนวิวัฒนาการของกระดูกโกลนจาก hyomandibula เป็นเหตุการณ์ต่างหากที่เกิดขึ้นก่อน
ทฤษฎี Reichert-Gaupp
โดยอาศัยไอเดียของนักธรรมชาติวิทยาชาวฝรั่งเศส Étienne Geoffroy Saint-Hilaire (พ.ศ. 2361) และของงานศึกษาต่าง ๆ รวมทั้งของนักกายวิภาคชาวเยอรมัน Johann Friedrich Meckel (2363), ของนักสรีรวิทยาชาวเยอรมัน Carl Gustav Carus (2361), ของนักคัพภวิทยาชาวเยอรมัน Martin Rathke (2368), และของนักวิทยาศาสตร์ชาวเอสโทเนีย (2371) นักคัพภวิทยาชาวเยอรมัน Karl Bogislaus Reichert จึงได้แสดงความสัมพันธ์ระหว่างขากรรไกรของสัตว์เลื้อยคลานกับกระดูกหูชั้นกลางของสัตว์เลี้ยงลูกด้วยนมในปี 2380 (ก่อนการพิมพ์หนังสือ On the Origin of Species ของดาร์วิน) ซึ่งต่อมาพัฒนาโดยนักกายวิภาคชาวเยอรมัน Ernst Gaupp จนกลายเป็นทฤษฎีที่เรียกว่า Reichert-Gaupp Theory
ในวิถีพัฒนาการของเอ็มบริโอ กระดูกทั่งและค้อนจะเกิดจาก first pharyngeal arch เหมือนกับ mandible และ maxilla และสัมพันธ์กับประสาทไทรเจมินัลสาขาเดียวกันด้วย
การค้นพบว่า กระดูกทั่งและค้อนของสัตว์เลี้ยงลูกด้วยนมจริง ๆ มีกำเนิดเดียวกับส่วนประกอบข้อต่อขากรรไกรของ "สัตว์เลื้อยคลาน" ... จัดเป็นหลักชัยอันหนึ่งในประวัติศาสตร์ของชีววิทยาเชิงเปรียบเทียบ
มันเป็นชัยชนะอันหนึ่งของโซ่งานวิจัยยาวเหยียดในสัตว์เลื้อยคลาน Theromorph ที่สูญพันธุ์ไปแล้ว เริ่มโดย Owen (2388) สืบต่อโดย Seeley, Broom, และ Watson ที่ได้แสดงขั้นตอนในระหว่าง ๆ ที่ความเปลี่ยนแปลงอาจเกิดจากกระดูก quadrate ข้างในไปเป็นข้อต่อ squamosal ข้างนอก ...
แต่ว่า การเปลี่ยนแปลงจากกระดูกขากรรไกรของ "สัตว์เลื้อยคลาน" ไปเป็นกระดูกหูชั้นกลางของ "สัตว์เลี้ยงลูกด้วยนม" ก็ไม่มีหลักฐานซากดึกดำบรรพ์เชื่อมจนกระทั่งคริสต์ทศวรรษ 1950 ที่ได้พบรายละเอียดในซากดึกดำบรรพ์ที่ปัจจุบันมีชื่อเสียงคือ Morganucodon
ยังมีงานวิจัยทางพันธุกรรมต่อมาเกี่ยวกับพัฒนาการของกระดูกหูจาก embryonic arch อีกด้วย และงานที่เชื่อมเรื่องนี้กับประวัติวิวัฒนาการ "ยีน Bapx1 หรือ Nkx3.2 ของสัตว์มีกระดูกสันหลัง มีกำเนิดเดียวกันกับยีน Bagpipe ของแมลงหวี่สกุล Drosophila เป็นสมาชิกในชั้น NK2 ของยีน homeobox..." งานวิจัยแสดงว่ายีนนี้เป็นเหตุในการเปลี่ยนกระดูกขากรรไกรที่พบในสัตว์อื่น ๆ ไปเป็นกระดูกหูในสัตว์เลี้ยงลูกด้วยนม ยีนอื่น ๆ ที่เกี่ยวข้องรวมทั้ง Dlx, Prx, และ Wnt

หูชั้นกลางมีจำกัดเฉพาะสัตว์เลี้ยงลูกด้วยนม
หูชั้นกลางของสัตว์เลี้ยงลูกด้วยนมมีกระดูกหูท่อนน้อย ๆ 3 ท่อนที่เรียกว่า กระดูกค้อน กระดูกทั่ง และกระดูกโกลน โดยกระดูกหูเป็นระบบคานงัดที่ซับซ้อน และมีหน้าที่ต่าง ๆ คือ ลดแอมพลิจูดของแรงสั่น เพิ่มแรงสั่น และดังนั้น ช่วยถ่ายทอดพลังงานเสียงอย่างมีประสิทธิภาพจากแก้วหูไปยังโครงสร้างของหูชั้นใน กระดูกหูทำงานทางแรงกลเหมือนกับหม้อแปลงไฟฟ้าทำกับกระแสไฟฟ้า เป็นการแมทชิงอิมพีแดนซ์ของแรงสั่นจากอากาศให้เป็นแรงสั่นในน้ำภายในคอเคลีย ซึ่งผลโดยรวมก็คือเพิ่มความไวและขีดจำกัดเสียงความถี่สูงของการได้ยินในสัตว์เลี้ยงลูกด้วยนมโดยเทียบกับสัตว์เลื้อยคลาน แต่รายละเอียดของโครงสร้างและผลเหล่านี้ก็ยังต่างกันอย่างเห็นได้ในสปีชีส์สัตว์เลี้ยงลูกด้วยนมต่าง ๆ แม้แต่สปีชีส์ที่ใกล้ชิดกันมากเช่นมนุษย์กับชิมแปนซี
ประวัติทางวิวัฒนาการ
นิยามที่จำกัด "สัตว์เลี้ยงลูกด้วยนม"
สปีชีส์สัตว์เลี้ยงลูกด้วยนมที่ยังมีอยู่สามารถระบุได้ด้วยต่อมน้ำนม (mammary gland) ในสัตว์ตัวเมีย แต่ว่า ลักษณะอื่นย่อมจำเป็นเพื่อกำหนดซากดึกดำบรรพ์ เพราะว่าต่อมน้ำนมและเนื้อเยื่ออื่น ๆ ไม่เหลืออยู่ในซากดึกดำบรรพ์ ดังนั้น นักบรรพชีวินวิทยาจึงต้องใช้ลักษณะเฉพาะที่มีในสัตว์เลี้ยงลูกด้วยนมที่ยังไม่สูญพันธุ์ทั้งหมด (รวมทั้งโมโนทรีม) แต่ไม่ปรากฏใน therapsid (สัตว์เลื้อยคลานคล้ายสัตว์เลี้ยงลูกด้วยนมซึ่งเป็นสัตว์บรรพบุรุษ) ในต้นยุคไทรแอสซิก ซึ่งก็คือสัตว์เลี้ยงลูกด้วยนมใช้กระดูกหูสองท่อนเพิ่มเพื่อการได้ยิน โดยเป็นกระดูกที่สัตว์มีถุงน้ำคร่ำ (Amniota) อื่น ๆ ใช้เพื่อกิน
สัตว์มีถุงน้ำคร่ำที่ไม่ใช่สัตว์เลี้ยงลูกด้วยนมทั้งหมดใช้ระบบกระดูกเช่นนี้รวมทั้ง กิ้งก่า จระเข้ ไดโนเสาร์ (และสัตว์ลูกหลานคือสัตว์ปีก) และ therapsid (บรรพบุรุษของสัตว์เลี้ยงลูกด้วยนม) ดังนั้น กระดูกหูชิ้นเดียวในหูชั้นกลางของสัตว์เหล่านี้ก็คือกระดูกโกลน
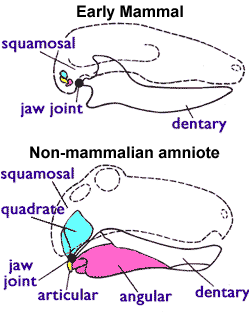
สัตว์เลี้ยงลูกด้วยนมมีข้อต่อขากรรไกรที่ต่างกัน ประกอบด้วยเพียงแค่ dentary (กระดูกขากรรไกรล่างซึ่งรองรับฟัน) และ squamosal (กระดูกกะโหลกศีรษะเล็กอีกชิ้นหนึ่ง) โดยกระดูก quadrate และ articular ที่เป็นส่วนของกระดูกขากรรไกรในสัตว์อื่น ได้วิวัฒนาการไปเป็นกระดูกค้อนและกระดูกทั่งในหูชั้นกลางเรียบร้อยแล้ว
ภาพรวมของหลักฐานดึกดำบรรพ์
นี่เป็นภาพต้นไม้แสดงสายพันธุ์ต่าง ๆ (ต้นไม้ชีวิต) แบบง่าย
--Tetrapod------
| (แปลตรง ๆ ว่า "มี 4 ขา" สัตว์ต้น ๆ หายใจผ่านเหงือก)
|
+-- สัตว์สะเทินน้ำสะเทินบก ----------------------------------------------
|
`--------Reptiliomorphs-----
| (สัตว์สะเทินน้ำสะเทินบก "คล้ายสัตว์เลื้อยคลาน")
|
`--สัตว์มีถุงน้ำคร่ำ------
|
+--Sauropsid (สัตว์ "หน้ากิ่งก่า") ---------------
| (กิ้งก่า จระเข้ ไดโนเสาร์ สัตว์ปีก
| เต่า และกลุ่มสัตว์ที่สูญพันธุ์แล้ว)
|
`--Synapsid------
|
`--Pelycosaur*----
|
`--Therapsid-----
|
`--สัตว์เลี้ยงลูกด้วยนม---------------
สัตว์มีกระดูกสันหลังที่เป็นสัตว์บกแท้พวกแรกคือ สัตว์มีถุงน้ำคร่ำ (amniote) ไข่ของพวกมันมีเยื่อข้างในที่ช่วยเก็บน้ำให้เอ็มบริโอที่กำลังเติบโตหายใจได้ ซึ่งทำให้สัตว์สามารถวางไข่บนบกแห้งได้ ในขณะที่สัตว์สะเทินน้ำสะเทินบกโดยทั่วไปจะต้องวางไข่ในน้ำ สัตว์มีถุงน้ำคร่ำปรากฏว่าเกิดในช่วงปลายยุคคาร์บอนิเฟอรัส จากสัตว์บรรพบุรุษ reptiliomorph (reptiliomorpha) ซึ่งเป็นกลุ่มสัตว์สะเทินน้ำสะเทินบกที่ลูกหลานที่ยังไม่สูญพันธุ์ล้วนเป็นสัตว์มีถุงน้ำคร่ำ ภายใน 2-3 ล้านปีต่อมา จึงเกิดสายพันธุ์สองสายที่ชัดเจน คือ synapsid ซึ่งเป็นบรรพบุรุษของสัตว์เลี้ยงลูกด้วยนม และ sauropsid ซึ่งเป็นบรรพบุรุษของกิ้งก่า งู จระเข้ ไดโนเสาร์ และสัตว์ปีก
ซากดึกดำบรรพ์เก่าแก่ที่สุดของกลุ่มสัตว์เหล่านี้มีอายุประมาณ 320-315 ล้านปีก่อน แต่โชคไม่ดีเพราะแน่ใจได้ยากว่าแต่ละกลุ่มวิวัฒนาการขึ้นมาเมื่อไร เพราะว่าซากดึกดำบรรพ์ของสัตว์มีกระดูกสันหลังจากปลายยุคคาร์บอนิเฟอรัสมีน้อยมาก และดังนั้น การเกิดขึ้นจริง ๆ ของสัตว์แต่ละกลุ่มอาจจะโบราณกว่าซากที่พบแล้ว
รูปแบบส่วนใหญ่ในหัวข้อต่อไปนี้ก็คือ กลุ่ม "ต่อมา" แต่ละกลุ่มจะเริ่มด้วยขากรรไกรและหูที่ "เก่าแก่" กว่าโดยได้จากบรรพบุรุษ แล้วจึงพัฒนามีขากรรไกรและหูที่ "ก้าวหน้า" กว่าต่อไป ให้สังเกตว่า ข้อต่อขากรรไกรและหูของสัตว์เลี้ยงลูกด้วยนม ไม่ได้วิวัฒนาการแบบก้าวต่อก้าวร่วมกับวิวัฒนาการลักษณะเฉพาะของสัตว์เลี้ยงลูกด้วยนมอื่น ๆ กล่าวอีกอย่างก็คือ ข้อต่อขากรรไกรและหูในช่วงต่าง ๆ ไม่ได้เป็นตัวระบุอะไรยกเว้นในระยะสุดท้าย ๆ ที่นักบรรพชีวินวิทยาเริ่มแยกว่าเป็นกายวิภาคโดยเฉพาะของสัตว์เลี้ยงลูกด้วยนม
หูของสัตว์สี่ขาและสัตว์มีถุงน้ำคร่ำต้น ๆ
ในสัตว์มีถุงน้ำคร่ำปัจจุบัน (รวมทั้งสัตว์เลี้ยงลูกด้วยนม) หูชั้นกลางรวบรวมเสียงจากอากาศผ่านแก้วหู แล้วถ่ายทอดแรงสั่นไปยังหูชั้นในผ่านโครงสร้างที่เป็นทั้งกระดูกอ่อนกระดูกแข็ง ซึ่งปกติจะรวมกระดูกโกลนด้วย แต่ว่า สัตว์สี่ขา สัตว์สะเทินน้ำสะเทินบก และสัตว์มีถุงน้ำคร่ำยุคต้น ๆ ที่สุด น่าจะไม่มีแก้วหู จริง ๆ แล้ว แก้วหูดูเหมือนจะวิวัฒนาการอย่างเป็นอิสระแล้ว ๆ เล่า ๆ 3-6 ครั้ง คือ
- ใน stegocephalian (สัตว์สะเทินน้ำสะเทินบกที่โบราณมาก)
- ใน anurans (กลุ่มสัตว์สะเทินน้ำสะเทินบกที่รวมกบและคางคก)
- ใน synapsid (รวมสัตว์เลี้ยงลูกด้วยนมและญาติ ๆ ที่สูญพันธุ์ไปแล้ว)
- ใน diapsid (กลุ่ม sauropsid ที่สำคัญที่สุดรวมทั้งกิ้งก่า จระเข้ ไดโนเสาร์ และสัตว์ปีก)
- อาจจะเกิดขึ้นต่างหากใน anapsids (เต่าและญาติที่สูญพันธุ์ไปแล้ว) ถ้าเต่าไม่ใช่ diapsid แปลง
- น่าจะใน seymouriamorph (กลุ่มหนึ่งของ reptiliomorph)
- และน่าจะใน temnospondyl (สัตว์สะเทินน้ำสะเทินบกโบราณ)
ในสัตว์ต้นสาย (basal) ทั้งหมดของ clade สัตว์มีถุงน้ำคร่ำใหญ่ ๆ ทั้งสาม (คือ synapsid, eureptile, และ parareptile) กระดูกโกลนเป็นส่วนค้ำจุนค่อนข้างใหญ่ของกระดูกหุ้มสมอง (braincase) และหน้าที่นี้ทำให้ไม่สามารถใช้ในระบบการได้ยินได้ แต่ก็มีหลักฐานเพิ่มขึ้นเรื่อย ๆ ว่า synapsid, eureptile และ parareptile ได้พัฒนาแก้วหูเชื่อมกับหูชั้นในผ่านกระดูกโกลนในช่วงยุคเพอร์เมียน
กระดูกขากรรไกรและหูของ therapsid ยุคต้น ๆ
ขากรรไกรของ synapsid ต้น ๆ รวมทั้งบรรพบุรุษของสัตว์เลี้ยงลูกด้วยนม ยังคล้ายกับของสัตว์สี่ขา (tetrapod) อื่น ๆ ในยุคเดียวกัน โดยขากรรไกรล่างมีกระดูก dentary ที่เป็นที่ยึดของฟัน และมีกระดูกเล็ก ๆ กว่าด้านหลังอีกหลายชิ้น ข้อต่อขากรรไกรประกอบด้วยกระดูก articular ในขากรรไกรล่างและ quadrate ในขากรรไกรบน
pelycosaur ต้น ๆ (ปลายยุคคาร์บอนิเฟอรัสและต้นยุคเพอร์เมียน) ไม่น่าจะมีแก้วหู และกระดูกโกลนที่ใหญ่มากทำหน้าที่ค้ำจุนกระดูกหุ้มสมอง (braincase) โดยมีส่วนล่างสุดวางอยู่บน quadrate
แต่ว่าสัตว์ลูกหลาน คือ therapsid (รวมทั้งบรรพบุรุษของสัตว์เลี้ยงลูกด้วยนม) น่าจะมีแก้วหูซึ่งอาจอยู่ติดกับกระดูก quadrate และกระดูกโกลนก็ยังอยู่ติดกับ quadrate แต่ทำหน้าที่เป็นกระดูกหูเพื่อการได้ยินไม่ใช่เพื่อค้ำจุนกระดูกหุ้มสมอง ดังนั้น quadrate ของ therapsid จึงมีหน้าที่คู่ คือเป็นทั้งส่วนของข้อต่อขากรรไกรและส่วนของระบบการได้ยิน


ขากรรไกรแบบข้อต่อคู่
ในช่วงยุคเพอร์เมียนและต้นยุคไทรแอสซิก กระดูก dentary ของ therapsid รวมทั้งบรรพบุรุษของสัตว์เลี้ยงลูกด้วยนม ใหญ่ขึ้นเรื่อย ๆ ในขณะที่กระดูกขากรรไกรอื่น ๆ เล็กลง ในที่สุด กระดูก dentary ก็กลายมาติดกับกระดูก squamosal ที่ขากรรไกรบนโดยอยู่ข้างหน้าของ quadrate รวมกันทั้งหมดเป็นข้อต่อของขากรรไกร 2 แบบพร้อม ๆ กัน คือ ข้อต่อด้านหน้าแบบสัตว์เลี้ยงลูกด้วยนม เป็นกระดูก dentary กับ squamosal และข้อต่อด้านหลังแบบสัตว์เลื้อยคลาน เป็น quadrate และ articular
ข้อต่อกระดูกขากรรไกรคู่เช่นนี้สามารถเห็นได้ในสัตว์ cynodont (therapsid ที่มี "ฟันสุนัข") หลัง ๆ และใน mammaliform (cynodont ที่มี "รูปร่างของสัตว์เลี้ยงลูกด้วยนม") ต้น ๆ สัตว์สกุล Morganucodon ที่มีชีวิตในช่วงปลายยุคไทรแอสซิกจนถึงกลางยุคจูแรสซิก (ที่ฟันดึกดำบรรพ์พบโดยมากในเทศมณฑล Glamorgan ในประเทศเวลส์) เป็น mammaliform แรก ๆ ที่พบและศึกษามากที่สุด เนื่องจากพบซากดึกดำบรรพ์มากเป็นพิเศษ คือ
Morganucodon เป็นสัตว์ระหว่างกลางที่เกือบสมบูรณ์ที่สุดในด้านนี้ (คือ มีขากรรไกรมีข้อต่อคู่) ระหว่างสัตว์เลื้อยคลานคล้ายสัตว์เลี้ยงลูกด้วยนม*ระดับสูงข้างหนึ่ง กับสัตว์เลี้ยงลูกด้วยนมธรรมดาอีกข้างหนึ่ง
- (* ให้สังเกตว่า "สัตว์เลื้อยคลานคล้ายสัตว์เลี้ยงลูกด้วยนม" เป็นคำเลิกใช้แล้วเพื่อเรียก therapsid)
กระดูกขากรรไกรและหูคล้ายสัตว์เลี้ยงลูกด้วยนม
ในขณะที่ dentary ขยายใหญ่ขึ้นเรื่อย ๆ ในยุคไทรแอสซิก ข้อต่อ quadrate-articular ที่เก่าแก่กว่าก็หมดประโยชน์ไปเรื่อย ๆ แม้ว่า กระดูกบางชิ้นจะหายไป แต่ quadrate (ซึ่งอยู่ต่อกับกระดูกโกลน), articular (ซึ่งต่อกับ quadrate), และ angular (ซึ่งต่อกับ articular) ก็กลายเป็นกระดูกอิสระที่สัมพันธ์กับกระดูกโกลน ซึ่งเกิดขึ้นอย่างน้อยสองครั้งใน mammaliformes ส่วนสัตว์กลุ่ม Multituberculates (สัตว์คล้ายสัตว์เลี้ยงลูกด้วยนมที่มีฟันเป็นตุ่ม ๆ) ซึ่งมีชีวิตตั้งแต่ 160 ล้านปีก่อน (กลางยุคจูแรสซิก) จนถึง 35 ล้านปีก่อน (ต้นสมัยโอลิโกซีน) มีข้อต่อขากรรไกรที่มีแต่กระดูก dentary และ squamosal โดยที่ quadrate และ articular ได้กลายเป็นส่วนของหูชั้นกลางแล้ว แต่ว่าลักษณะต่าง ๆ ของฟัน ขากรรไกร และกะโหลกศีรษะก็ยังต่างจากสัตว์เลี้ยงลูกด้วยนมอย่างสำคัญ
| ความสัมพันธ์ของต้นสายพันธุ์สัตว์เลี้ยงลูกด้วยนมกับกลุ่มอื่น ๆ | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
ในสายพันธุ์ที่ใกล้ชิดกับสัตว์เลี้ยงลูกด้วยนมมากที่สุด ขากรรไกรของ Hadrocodium (ประมาณ 195 ล้านปีก่อนในยุคจูแรสซิกต้น ๆ) แสดงนัยว่า สัตว์หรือว่าบรรพบุรุษใกล้ ๆ อาจเป็นสัตว์พวกแรกที่มีหูชั้นกลางเหมือนกับสัตว์เลี้ยงลูกด้วยนมแทบสมบูรณ์ คือ มันไม่มีร่องที่ด้านหลังของขากรรไกรล่าง ซึ่งเป็นที่ยึดแก้วหูของ therapsid และ mammaliformes ก่อน ๆ การไม่มีร่องเช่นนี้แสดงนัยว่า หูของ Hadrocodium เป็นส่วนของกระดูกหุ้มสมอง เหมือนกับสัตว์เลี้ยงลูกด้วยนม และดังนั้น กระดูกเดิมที่เป็น articular และ quadrate ได้กลายเป็นส่วนของหูชั้นกลาง คือได้กลายเป็นกระดูกค้อนและกระดูกทั่งแล้ว แต่ว่ากระดูก dentary ก็ยังมีช่องเว้าที่ด้านหลังซึ่งสัตว์เลี้ยงลูกด้วยนมไม่มี ซึ่งแสดงนัยว่า dentary อาจมีรูปร่างแบบนี้ถ้า articular และ quadrate ยังเป็นส่วนของข้อต่อกระดูกขากรรไกร
มีการเสนอในปี 2548 ว่า ช่องเว้าที่ค่อนข้างใหญ่ของกระดูกขากรรไกรของโมโนทรีมต้นยุคครีเทเชียสสกุล Teinolophos เป็นหลักฐานข้อต่อขากรรไกรของบรรพบุรุษสัตว์เลี้ยงลูกด้วยนม (pre-mammalian) คือเพราะว่า therapsid และ mammaliform หลายชนิดก็มีช่องเว้านี้เหมือนกัน ซึ่งเป็นจุดต่อกับกระดูก articular กับ angular และดังนั้น Teinolophos จึงมีหูชั้นกลางเหมือนกับบรรพบุรุษสัตว์เลี้ยงลูกด้วยนม และดังนั้น วิวัฒนาการของกระดูกหูของหูชั้นกลางแบบสัตว์เลี้ยงลูกด้วยนมของโมโนทรีม จึงเกิดขึ้นอย่างเป็นอิสระจากสัตว์เลี้ยงลูกด้วยนมอื่น ๆ
แต่การวิเคราะห์ปี 2551 สรุปว่า Teinolophos เป็นตุ่นปากเป็ดอย่างสมบูรณ์ และส่วนเว้านั้นเป็นช่องสำหรับเส้นประสาทจำนวนมากที่มาจากตัวรับกระแสไฟฟ้าและแรงสั่นที่ปาก (ซึ่งเป็นลักษณะเฉพาะของตุ่นปากเป็ดในบรรดาโมโนทรีม) และดังนั้น รอยเว้าจึงไม่ใช่เป็นหลักฐานว่า Teinolophos มีข้อต่อขากรรไกรและหูชั้นกลางเหมือนกับบรรพบุรุษสัตว์เลี้ยงลูกด้วยนม แต่ก็แปลกดีว่า ผู้เขียน 2 ท่านในงานปี 2548 ก็เป็นผู้เขียนงานปี 2551 ด้วย แถมงานหลังก็คัดค้านงานแรก
ซากดึกดำบรรพ์ช่วงเปลี่ยนสภาพที่ค้นพบในปี 2550 เป็นสัตว์เลี้ยงลูกด้วยนมยุคต้น ๆ สกุล Yanoconodon จาก 125 ล้านปีก่อนในมหายุคมีโซโซอิก ที่กระดูกหูทั้งสามได้แยกออกจากขากรรไกรแล้วทำหน้าที่ทางการได้ยินในหูชั้นกลาง แต่กระดูกก็ยังเชื่อมกับขากรรไกรผ่านกระดูกที่เรียกว่า Meckel's cartilage แต่โดยเป็นกระดูกแข็ง ซึ่งในสัตว์เลี้ยงลูกด้วยนมต่อ ๆ มาจะเป็นกระดูกอ่อนที่หายไปในช่วงพัฒนาการ
ผลต่อการได้ยิน
พิสัยความถี่และความไวเสียงของหูจะขึ้นอยู่กับรูปร่างและระเบียบของกระดูกหูชั้นกลาง ใน synapsid ต้น ๆ เช่น pelycosaur กระดูก quadrate และ articular เป็นข้อต่อขากรรไกร ซึ่งจำกัดการใช้กระดูกเหล่านี้เพื่อเปลี่ยนพิสัยความถี่เสียงของหู แต่เมื่อกระดูกเหล่านี้ไม่เป็นส่วนของกระดูกขากรรไกรแล้ว การเปลี่ยนแปลงกระดูกเพื่อการได้ยินก็จะไม่มีผลต่อการทำงานของขากรรไกร และนี่ทำให้สามารถเกิดวิวัฒนาการของกระดูกอย่างไม่จำกัดในสัตว์เลี้ยงลูกด้วยนม และโดยยุคจูแรสซิก หูที่เป็นลักษณะเฉพาะของสัตว์เลี้ยงลูกด้วยนมก็เริ่มปรากฏแล้ว โดยที่กระดูก angular ได้กลายมาเป็น tympanic annula (ซึ่งเป็นกระดูกค้ำจุนแก้วหู) ในขณะที่ articular และ quadrate ได้กลายมาเป็นกระดูกค้อนและกระดูกทั่งตามลำดับ โดยเชื่อมเป็นลูกโซ่กับกระดูกโกลน กระดูกหูทั้งสามทำหน้าที่เป็นระบบจับคู่อิมพีแดนซ์ เพื่อปรับปรุงการถ่ายทอดเสียงเพื่อให้ได้ยินดีขึ้น
การเปลี่ยนสภาพของกระดูกหูเหล่านี้มีหลักฐานดีที่สุด ในเรื่องทางวิวัฒนาการทั้งหมด และซากดึกดำบรรพ์ที่ค้นพบใหม่ ๆ และมาจากยุคเปลี่ยนสภาพ ก็ได้ปรับปรุงความเข้าใจในเรื่องนี้ให้ดีขึ้น แต่ก็แสดงด้วยว่า นี่ไม่ใช่กระบวนการเปลี่ยนสภาพแบบเป็นเส้นตรงจากขากรรไกร (ข้อต่อแบบ quadrate-articular) และหูชั้นกลาง (กระดูกโกลนเป็นกระดูกหูเดียว) ของ therapsid ต้น ๆ มาสู่กายวิภาคหูของสัตว์เลี้ยงลูกด้วยนมปัจจุบัน
การคัดเลือกโดยธรรมชาติ
มีการเสนอว่า การคัดเลือกโดยธรรมชาติอาจเป็นปัจจัยผลักดันโครงสร้างหูชั้นกลางของสัตว์เลี้ยงลูกด้วยนม คือ สัตว์เลี้ยงลูกด้วยนมยุคต้น ๆ หลายอย่างค่อนข้างเล็ก และฟันแสดงว่า พวกมันเป็นสัตว์กินแมลง ถ้าพวกมันเป็นสัตว์เลือดอุ่นเหมือนกับสัตว์เลี้ยงลูกด้วยนมปัจจุบัน ก็อาจเป็นสัตว์หากินกลางคืนด้วย ซึ่งเข้ากับจินตภาพยอดนิยมของสัตว์เลี้ยงลูกด้วยนมว่าเป็นสัตว์เล็ก ๆ หากินแมลงเวลากลางคืน และมีชีวิตรอดในระบบนิเวศเฉพาะ ๆ ที่ไดโนเสาร์ใหญ่ผู้ครองโลกในยุคเดียวกันเข้าไม่ถึง การได้ยินเสียงที่ดีกว่า โดยเฉพาะที่ความถี่สูง ก็จะเป็นประโยชน์ต่อสัตว์หากินกลางคืน โดยเฉพาะเพื่อตรวจจับหาแมลง สถานการณ์เยี่ยงนี้ เข้ากับความได้เปรียบโดยการคัดเลือกอันเป็นปัจจัยเสริมให้เกิดการเปลี่ยนสภาพ
ประวัติและภาพรวม
ถ้าอนุมานนอกช่วงการเกิดสัณฐานช่วงพัฒนาการ (developmental morphogenesis) จากงานศึกษาทางพันธุกรรม ให้เข้ากับหลักฐานซากดึกดำบรรพ์ของสัตว์เลี้ยงลูกด้วยนมต้น ๆ วิวัฒนาการของหูชั้นกลางในสัตว์เลี้ยงลูกด้วยนมต้น ๆ ก็จะเป็นกรณีศึกษาแบบบูรณาการที่แสดงว่า พัฒนาการสามารถมีผลเชิงกลต่อการเปลี่ยนรูปของคอมเพล็กซ์โครงสร้างสำคัญในกระบวนการวิวัฒนาการได้อย่างไร
— Zhe-Xi Luo (2011). Developmental Patterns in Mesozoic Evolution of Mammal Ears
แม้ว่ากระดูกโกลนจะมีอยู่ในสัตว์สี่ขาหลายอย่าง แต่การเพิ่มทั้งกระดูกค้อนและกระดูกทั่ง (หรือ quadrate และ articular) ในหูชั้นกลางอันเป็นลักษณะเฉพาะของกลุ่มสัตว์เลี้ยงลูกด้วยนม ได้แยกกลุ่มออกจากสัตว์เลื้อยคลานและสัตว์มีกระดูกสันหลังอื่น ๆ ทั้งหมด ในอดีตเวลาหนึ่ง ต้นไม้ชีวิตจึงได้ปรากฏเหมือนกับไม่สืบตอน
แต่โดยต้นคริสต์ศตวรรษที่ 19 ก็เกิดสมมติฐานว่า กระดูกเหล่านี้ไม่ใช่อะไรใหม่โดยสิ้นเชิง แต่เป็นกระดูก 2 ชิ้นในขากรรไกรของสัตว์ที่ไม่ใช่สัตว์เลี้ยงลูกด้วยนมอื่น ๆ สมมติฐานนี้มีเหตุผล และไม่ใช่เพราะการมีกระดูกหูสามชิ้นเท่านั้น แต่เพราะลักษณะทางกายวิภาคอื่น ๆ เช่น เส้นทางของเส้นประสาทในศีรษะ
เมื่อสาขาชีววิทยาเชิงวิวัฒนาการเริ่มขยายตัว จึงได้ปฏิบัติต่อความสัมพันธ์เช่นนี้เหมือนว่า สัตว์เหล่านี้ (สัตว์เลื้อยคลาน สัตว์เลี้ยงลูกด้วยนม และสัตว์มีกระดูกสันหลังอื่น ๆ) สืบเชื้อสายร่วมกัน เพื่อที่คำอธิบายทางวิวัฒนาการจะสมเหตุผล กระดูกจึงต้องเปลี่ยนหน้าที่จากเป็นส่วนของกลไกการกินอาหารคือข้อต่อขากรรไกร ไปเป็นส่วนของการได้ยินเท่านั้น ซึ่งหมายความว่า จะต้องมีซากดึกดำบรรพ์ที่แสดงสภาพในระหว่าง คือแสดงการสืบต่อของสองหน้าที่นี้
การค้นพบซากดึกดำบรรพ์ของ Morganucodon และอื่น ๆ จึงทำให้มีตัวอย่างที่เป็นรูปธรรมในเรื่องนี้ เพราะว่า ซากที่พบมีข้อต่อขากรรไกรทั้งสองอย่าง คือ แบบ "สัตว์เลื้อยคลานเก่า" และแบบ "สัตว์เลี้ยงลูกด้วยนมใหม่" เป็นหลักฐานยืนยันรูปแบบการอนุมานจากสาขากายวิภาคเปรียบเทียบไปยังสาขาชีววิทยาเชิงวิวัฒนาการ (ดูคำพูดของ Zhe-Xi Luo ด้านบน)
สัตว์เลี้ยงลูกด้วยนมต้น ๆ โดยทั่วไปเป็นสัตว์เล็ก ๆ และน่าจะเป็นสัตว์หากินแมลงเวลากลางคืน ทฤษฎีนี้ใช้อธิบายกลไกทางวิวัฒนาการที่ขับเคลื่อนความเปลี่ยนแปลงได้ เพราะด้วยกระดูกหูเล็ก ๆ ในหูชั้นกลาง สัตว์เลี้ยงลูกด้วยนมจึงสามารถขยายพิสัยความถี่เสียงที่ได้ยิน ซึ่งช่วยตรวจจับหาแมลงในที่มืด ทฤษฎีการคัดเลือกโดยธรรมชาติก็จะสามารถอธิบายชัยชนะของคุณลักษณะเยี่ยงนี้
แหล่งข้อมูลอื่น
-
Allin, EF; Hopson, JA (1992). "Chapter 28: Evolution of the Auditory System in Synapsida ("Mammal-Like Reptiles" and Primitive Mammals) as Seen in the Fossil Record". The Evolutionary biology of hearing. Berlin: Springer-Verlag. pp. 587–614. ISBN 0-387-97588-8.
{{cite book}}: ไม่รู้จักพารามิเตอร์|editors=ถูกละเว้น แนะนำ (|editor=) (help)CS1 maint: uses authors parameter (ลิงก์) - Anthwal, Neal; Joshi, Leena; Tucker, Abigail S (2012). "Evolution of the mammalian middle ear and jaw: adaptations and novel structures". Journal of Anatomy. 222 (1): 1–96. doi:10.1111/j.1469-7580.2012.01526.x. PMC 3552421. PMID 22686855.
- Arthur, Wallace (2011). "10.3 Compound Repatterning at a Single Level of Organisation". Evolution: A developmental approach. Oxford: Wiley-Blackwell. pp. 151–155. ISBN 978-1-4051-8658-2.
- Asher, Robert J. (2012). Evolution and belief: confessions of a religious paleontologist. Cambridge & New York: Cambridge University Press. pp. 93–110, 196–200. ISBN 9780521193832.
- Gould, SJ (1993). "Chapter 6: An Earful of Jaw". Eight Little Piggies: reflections in natural history. New York: Norton. ISBN 0-393-03416-X.
-
Hopson, JA (1987-01). "The mammal-like reptiles: a study of transitional fossils". The American Biology Teacher. 49 (1): 16–26. doi:10.2307/4448410. JSTOR 4448410.
{{cite journal}}: ตรวจสอบค่าวันที่ใน:|date=(help) - Kielan-Jaworowska, Z (2013). "5. Origins of Mammals and the Earliest Representatives of Mammaliforms and Mammals". In Pursuit of Early Mammals. Life of the Past. Bloomington, Indiana: Indiana University Press. pp. 73–96. ISBN 978-0-253-00824-4. especially pages 85-96
- Luo, ZX; Kielan-Jaworowska, Z; Cifelli, RL (2004). "Chapter 3: Origin of mammals". Mammals from the age of dinosaurs: origins, evolution, and structure. New York: Columbia University Press. ISBN 0-231-11918-6.
-
Manley, GA; Sienknecht, UJ (2013). "Chapter 2: The Evolution and Development of Middle Ears in Land Vertebrates". The Middle Ear: Science, Otosurgery, and Technology. Springer Handbook of Auditory Research. Vol. 46. New York: Springer. pp. 7–30. doi:10.1007/978-1-4614-6591-1_2. ISBN 978-1-4614-6590-4.
{{cite book}}: ไม่รู้จักพารามิเตอร์|editors=ถูกละเว้น แนะนำ (|editor=) (help)CS1 maint: uses authors parameter (ลิงก์) -
Meng, J; Zheng, XT; Wang, XL (2016). "Ear Ossicle Morphology of the Jurassic Euharamiyidan Arboroharamiya and Evolution of Mammalian Middle Ear". Journal of Morphology. doi:10.1002/jmor.20565.
{{cite journal}}: CS1 maint: uses authors parameter (ลิงก์) contains wide bibliography of scientific literature up to 2016 -
Rosowski, JJ (1992). "Chapter 29: Hearing in Transitional Mammals: Predictions from the Middle-Ear Anatomy and Hearing Capabilities of Extant Mammals". The Evolutionary biology of hearing. Berlin: Springer-Verlag. pp. 615–632. ISBN 0-387-97588-8.
{{cite book}}: ไม่รู้จักพารามิเตอร์|editors=ถูกละเว้น แนะนำ (|editor=) (help) -
Rougier, GW; White, JR (2006). "Chapter 6: Major Changes in the Ear Region and Basicranium of Early Mammals". Amniote paleobiology: perspectives on the evolution of mammals, birds, and reptiles: a volume honoring James Allen Hopson. Chicago: University of Chicago Press. pp. 269–311. ISBN 0-226-09477-4.
{{cite book}}: ไม่รู้จักพารามิเตอร์|editors=ถูกละเว้น แนะนำ (|editor=) (help)CS1 maint: uses authors parameter (ลิงก์) - Shubin, N (2008). "Chapter 10: Ears". Your inner fish: a journey into the 3.5-billion-year history of the human body. New York: Pantheon Books. ISBN 0-375-42447-4.
- Tucker, Abigail S. (2017-02-05). "Major evolutionary transitions and innovations: the tympanic middle ear" (PDF). Philosophical Transactions of the Royal Society B. 372 (1713). doi:10.1098/rstb.2015.0483.
- Theobald, D (2004). "29+ Evidences for Macroevolution: Part 1, Example 2: reptile-mammals". TalkOrigins. สืบค้นเมื่อ 2009-06-17.
- "The Evolution of Hearing from Amphibians to Mammals". 2004. คลังข้อมูลเก่าเก็บจากแหล่งเดิมเมื่อ 2015-11-14. สืบค้นเมื่อ 2016-03-26.
- Cuffey, CA (2001). "The Fossil Record: Evolution or "Scientific Creation": Mammal-Like Reptiles". GCSSEPM Foundation. คลังข้อมูลเก่าเก็บจากแหล่งเดิมเมื่อ 2009-05-01. สืบค้นเมื่อ 2009-06-17.
-
Matzke, N (2005). "The testimony of Kevin Padian in Kitzmiller v. Dover". sciohost.org. คลังข้อมูลเก่าเก็บจากแหล่งเดิมเมื่อ 2009-04-14. สืบค้นเมื่อ 2009-06-17.
Based on testimony by Kevin Padian in the case of Kitzmiller v. Dover
- Your Inner Fish : We Hear With the Bones That Reptiles Eat With
| วิวัฒนาการ |  |
|
|---|---|---|
| พันธุศาสตร์ประชากร | ||
| พัฒนาการ | ||
| วิวัฒนาการของอนุกรมวิธาน | ||
| วิวัฒนาการของอวัยวะ | ||
| วิวัฒนาการของชีวกระบวนการ | ||
| กระแสชีวิตและแบบจำลอง | ||
| แบบจำลองสายพันธุ์ใหม่ | ||
| บทความที่เกี่ยวข้อง | ||
| ประวัติชีวศาสตร์ | ||