
| (Neuron) | |
|---|---|

ภาพวาดเซลล์ประสาทของสมองน้อยนกพิราบปี พ.ศ. 2442 โดยนักประสาทวิทยาศาสตร์ชาวสเปน นพ. ซานเตียโก รามอน อี กาฆัล (A) เป็นเซลล์เพอร์คินจี และ (B) เป็น granule cells โดยทั้งสองแบบเป็นเซลล์ประสาทหลายขั้ว (multipolar)
| |
| ตัวระบุ | |
| MeSH | D009474 |
| นิวโรเล็กซ์ ID | sao1417703748 |
| TA98 | A14.0.00.002 |
| TH | H2.00.06.1.00002 |
| FMA | 54527 |
| ศัพท์ทางกายวิภาคของประสาทกายวิภาคศาสตร์ | |
เซลล์ประสาท หรือ นิวรอน (อังกฤษ: neuron, /ˈnjʊərɒn/ nyewr-on, หรือ /ˈnʊərɒn/ newr-on) เป็นเซลล์เร้าได้ด้วยกระแสไฟฟ้าที่ทำหน้าที่ประมวลและส่งข้อมูลผ่านสัญญาณไฟฟ้าและเคมี โดยส่งผ่านจุดประสานประสาท (synapse) ซึ่งเป็นการเชื่อมต่อโดยเฉพาะกับเซลล์อื่น ๆ นิวรอนอาจเชื่อมกันเป็นโครงข่ายประสาท (neural network) และเป็นองค์ประกอบหลักของสมองกับไขสันหลังในระบบประสาทกลาง (CNS) และของปมประสาท (ganglia) ในระบบประสาทนอกส่วนกลาง (PNS) นิวรอนที่ทำหน้าที่โดยเฉพาะ ๆ รวมทั้ง
- เซลล์ประสาทรับความรู้สึก (sensory neuron) ตอบสนองต่อสัมผัส เสียง แสง และสิ่งเร้าอื่น ๆ แล้วส่งต่อสัญญาณ/ข้อมูลไปยังไขสันหลังและสมองต่อไป
- เซลล์ประสาทสั่งการ (motor neuron) รับสัญญาณจากสมองและไขสันหลัง แล้วทำให้กล้ามเนื้อเกร็งคลาย และทำให้ต่อมต่าง ๆ หลั่งสาร
- เซลล์ประสาทต่อประสาน (interneuron) เชื่อมนิวรอนต่าง ๆ ในสมองเขตเดียวกัน หรือเชื่อมเป็นโครงข่ายประสาทในไขสันหลัง
นิวรอนโดยทั่วไปจะมีตัวเซลล์ที่เรียกว่า soma, ใยประสาทนำเข้า/เดนไดรต์ (dendrite), และแกนประสาทนำออก/แอกซอน (axon) คำภาษาอังกฤษว่า neurite สามารถใช้เรียกทั้งเดนไดรต์และแอกซอน โดยเฉพาะในระยะที่เซลล์ยังไม่พัฒนาแยกเป็นเซลล์โดยเฉพาะ ๆ (undifferentiated) ส่วนเดนไดรต์เป็นโครงสร้างบาง ๆ ที่ยื่นออกจากตัวเซลล์ บ่อยครั้งยาวเป็นร้อย ๆ ไมโครเมตรและแยกออกเป็นสาขา ๆ รวมกันเป็นต้นไม้เดนไดรต์ (dendritic tree) ที่ซับซ้อน แอกซอน (หรือเรียกว่า nerve fiber ถ้ามีปลอกไมอีลิน) เป็นโครงสร้าง (process) ที่ยื่นออกจากตัวเซลล์ตรงตำแหน่งเฉพาะที่เรียกว่า axon hillock และอาจยาวถึง 1 เมตรในมนุษย์หรือมากกว่านั้นในสปีชีส์อื่น ๆ "nerve fiber" บ่อยครั้งรวมตัวกันเป็น "nerve fascicle" และในระบบประสาทนอกส่วนกลาง (PNS) fascicle จะรวมตัวกันเป็น "nerve" แม้ว่าตัวเซลล์ประสาทเองอาจจะมีเดนไดรต์ยื่นออกหลายอัน แต่จะมีแอกซอนยื่นออกเพียงแค่อันเดียว ถึงกระนั้น แอกซอนก็อาจแตกสาขาเป็นร้อย ๆ ครั้งก่อนจะสิ้นสุดลง
ในจุดประสานประสาทโดยมาก สัญญาณจะดำเนินจากแอกซอนของเซลล์หนึ่ง ไปยังเดนไดรต์ของอีกเซลล์หนึ่ง แต่ว่า ก็มีข้อยกเว้นมากมาย เช่น นิวรอนอาจไร้เดนไดรต์ หรือไร้แอกซอน และจุดประสานประสาทสามารถเชื่อมแอกซอนกับแอกซอน หรือเชื่อมเดนไดรต์กับเดนไดรต์
นิวรอนทั้งหมดสามารถเร้าได้โดยกระแสไฟฟ้า โดยรักษาศักย์ไฟฟ้าที่ต่างกันระหว่างภายในภายนอกเซลล์ข้ามเยื่อหุ้มเซลล์ โดยใช้ปั๊มไอออน (หรือ ion transporter) บวกกับช่องไอออนที่ฝังอยู่ในเยื่อหุ้มเซลล์เพื่อรักษาความเข้มข้นของไอออนต่าง ๆ (เช่น โซเดียม โพแทสเซียม คลอไรด์ และแคลเซียม) ในระดับที่ต่างกันระหว่างภายในภายนอกเซลล์ ความเปลี่ยนแปลงของความต่างศักย์ไฟฟ้าข้ามเยื่อหุ้มเซลล์อาจเปลี่ยนการทำงานของช่องไอออนที่เปิดปิดโดยศักย์ไฟฟ้า (Voltage-gated ion channel) ถ้าศักย์ต่างเปลี่ยนมากพอ ก็จะมีผลเป็นศักยะงาน (action potential) ที่ยิงแบบเกิดหรือไม่เกิด (all-or-none) และเป็นพัลส์ไฟฟ้าเคมีที่วิ่งไปอย่างรวดเร็วทางแอกซอนของเซลล์ แล้วจบลงด้วยการส่งสัญญาณข้ามเซลล์ที่จุดประสานประสาท (โดยเซลล์ประสาทต่อไปอาจส่งสัญญาณต่อ)
ในกรณีโดยมาก นิวรอนเกิดจากเซลล์ต้นกำเนิดโดยเฉพาะ ๆ และนิวรอนในสมองผู้ใหญ่ปกติจะไม่มีการแบ่งเซลล์ แต่ก็พบว่า astrocyte ซึ่งเป็นเซลล์เกลีย (glial cell) รูปดาว สามารถเปลี่ยนเป็นนิวรอนได้เพราะมีลักษณะ pluripotency ของเซลล์ต้นกำเนิด กำเนิดของเซลล์ประสาท (Neurogenesis) โดยมากในสมองจะหยุดลงเมื่อถึงวัยผู้ใหญ่ แต่ก็มีหลักฐานที่ชัดเจนว่ามีนิวรอนเกิดใหม่จำนวนมากในเขตสมองสองเขต คือที่ฮิปโปแคมปัส และที่ olfactory bulb ซึ่งเป็นโครงสร้างประสาทในสมองส่วนหน้าที่มีหน้าที่เกี่ยวกับการได้กลิ่น
| เซลล์ประสาทของระบบประสาทนอกส่วนกลาง (PNS) |
|---|


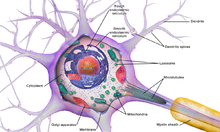
ภาพรวม
นิวรอนเป็นเซลล์โดยเฉพาะที่พบในสัตว์กลุ่ม eumetozoa ทุกอย่าง และมีสัตว์เพียงบางอย่างเช่น ฟองน้ำ และสัตว์ไม่ซับซ้อนบางประเภท ที่ไร้นิวรอน
ลักษณะที่เป็นตัวกำหนดนิวรอนก็คือสามารถเร้าได้ด้วยกระแสไฟฟ้า และการมีจุดประสานประสาท (synapse) ซึ่งเป็นจุดต่อซับซ้อนของเยื่อหุ้มเซลล์ที่ส่งสัญญาณไปยังเซลล์อื่น ๆ
นิวรอนกับเซลล์เกลียที่เป็นตัวสนับสนุนโดยโครงสร้างและเมแทบอลิซึม รวมกันเป็นระบบประสาท ในสัตว์มีกระดูกสันหลัง นิวรอนส่วนมากจะอยู่ในระบบประสาทกลาง (CNS) แต่ก็มีบางส่วนอยู่ที่ปมประสาท (ganglia) ในระบบประสาทนอกส่วนกลาง (PNS) และมีเซลล์ประสาทรับความรู้สึกจำนวนมากในอวัยวะรับความรู้สึกเช่นจอตาและหูชั้นในรูปหอยโข่ง (cochlea)
นิวรอนปกติจะแบ่งเป็น 3 ส่วน คือ ตัวเซลล์ (soma หรือ cell body) ใยประสาทนำเข้า/เดนไดรต์ (dendrite) และแกนประสาทนำออก/แอกซอน (axon) ตัวเซลล์ปกติจะมีลักษณะกระชับ แต่เดนไดรต์และแอกซอนจะยื่นออกไปจากตัวเซลล์ เดนไดรต์ปกติจะมีสาขามากมาย โดยแต่ละสาขาจะบางลง ๆ และจะยื่นออกไปไกลที่สุด 2-3 ร้อยไมโครเมตรจากตัวเซลล์ ส่วนแอกซอนจะยื่นออกจากตัวเซลล์ที่ axon hillock และสามารถไปได้ไกล โดยมีสาขาเป็นร้อย ๆ แต่ว่าไม่เหมือนกับเดนไดรต์ แอกซอนจะดำรงขนาดไปจนสุด ตัวเซลล์อาจจะมีเดนไดรต์มากมายแต่จะมีแอกซอนเพียงแค่อันเดียว
ตัวเซลล์และเดนไดรต์เป็นตัวรับสัญญาณไซแนปส์จากเซลล์ประสาทอื่น ๆ ส่วนสัญญาณจะส่งผ่านแอกซอน ดังนั้น จุดประสานประสาทโดยทั่วไปก็คือการเชื่อมต่อกันระหว่างแอกซอนของเซลล์หนึ่งไปยังเดนไดรต์หรือตัวเซลล์ของนิวรอนอีกตัวหนึ่ง สัญญาณที่ส่งอาจจะเป็นแบบเร้า (excitatory) หรือแบบห้าม (inhibitory) ถ้าการเร้าสุทธิที่นิวรอนได้รับในระยะเวลาสั้น ๆ มีระดับเพียงพอ นิวรอนก็จะสร้างพัลส์ที่เรียกว่าศักยะงาน (action potential) เริ่มที่ตัวเซลล์แล้วส่งไปอย่างรวดเร็วทางแอกซอน เมื่อถึงที่สุด ปลายนิวรอน (axon terminal) ก็จะส่งสัญญาณต่อที่จุดประสานประสาทไปยังนิวรอนอื่น ๆ ที่เชื่อมต่อ
นิวรอนจำนวนมากมีลักษณะดังที่ว่า แต่ว่าก็มีข้อยกเว้นทุกอย่าง คือ มีทั้งนิวรอนที่ไม่มีตัวเซลล์ ไม่มีเดนไดรต์ และไม่มีแอกซอน นอกจากจุดประสานประสาทที่ส่งสัญญาณไปยังเดนไดรต์ (axodendritic) หรือตัวเซลล์ (axosomatic) แล้ว ยังมีจุดประสานที่ส่งสัญญาณจาก แอกซอนไปยังแอกซอน (axoaxonic, axon-to-axon) หรือจากเดนไดรต์ไปยังเดนไดรต์ (dendrodendritic, dendrite-to-dendrite)
หน้าที่หลักของนิวรอนก็คือการส่งสัญญาณข้ามจุดประสานประสาท ซึ่งโดยส่วนหนึ่งเป็นทางไฟฟ้าและส่วนหนึ่งเป็นทางเคมี คุณลักษณะทางไฟฟ้าจะขึ้นอยู่กับคุณสมบัติของเยื่อหุ้มเซลล์ประสาท
เหมือนกับเซลล์อื่น ๆ ในสัตว์ ตัวเซลล์ของนิวรอนจะหุ้มด้วยเยื่อหุ้มเซลล์ ซึ่งเป็นโมเลกุลลิพิด 2 ชั้นที่มีโครงสร้างโปรตีนต่าง ๆ ฝังอยู่ เยื่อลิพิด 2 ชั้นเช่นนี้เป็นฉนวนไฟฟ้าที่ดี แต่ในนิวรอน โครงสร้างโปรตีนที่ฝังอยู่ในเยื่อจะมีฤทธิ์ทางไฟฟ้า รวมทั้งช่องไอออนที่เปิดให้ไอออนขั้วบวกหรือลบข้ามผ่านเยื่อ และปั๊มไออนที่ปั๊มไอออนจากด้านหนึ่งของเยื่อไปยังอีกด้านหนึ่ง
ช่องไอออนโดยมากเปิดให้ไอออนโดยเฉพาะ ๆ ข้าม บางอย่างเปิดปิดโดยศักย์ไฟฟ้า (voltage gated) ซึ่งหมายความว่าจะอยู่ในสภาพเปิดปิดขึ้นอยู่กับความแตกต่างของศักย์ไฟฟ้าข้ามเยื่อ บางอย่างเปิดปิดทางเคมี (chemically gated) ซึ่งหมายความว่าจะอยู่ในสภาพเปิดปิดโดยอาศัยปฏิสัมพันธ์ทางเคมีกับของเหลวที่อยู่ระหว่างเซลล์
ปฏิสัมพันธ์ของช่องไอออนและปั๊มไอออนทำให้เกิดความต่างศักย์ข้ามเยื่อหุ้มเซลล์ โดยปกติอยู่ที่ประมาณน้อยกว่า 1/10 โวลต์เล็กน้อย ศักย์ไฟฟ้าที่ว่ามีหน้าที่สองอย่าง อย่างแรกคือเป็นแหล่งพลังงานสำหรับโครงสร้างโปรตีนที่เปิดปิดโดยศักย์ไฟฟ้าที่ฝังอยู่ในเยื่อ อย่างที่สองคือเป็นมูลฐานการส่งสัญญาณไฟฟ้าระหว่างส่วนต่าง ๆ ของเยื่อหุ้มเซลล์
นิวรอนส่งสัญญาณผ่านจุดประสานประสาททางเคมี (chemical synapse) และจุดประสานประสาททางไฟฟ้า (electrical synapse) ในกระบวนการที่เรียกว่าการสื่อประสาท (neurotransmission) หรือการสื่อผ่านไซแนปส์ (synaptic transmission) กระบวนการหลักที่จุดชนวนให้เซลล์ปล่อยสารสื่อประสาท (neurotransmitter) ก็คือศักยะงาน (action potential) ซึ่งเป็นสัญญาณไฟฟ้าวิ่งผ่าน และสร้างโดยอาศัยความต่างศักย์ของเยื่อหุ้มเซลล์/เยื่อหุ้มเซลล์ที่เร้าได้ด้วยไฟฟ้า โดยเกิดในลักษณะเป็นคลื่นของการลดขั้ว (depolarization)
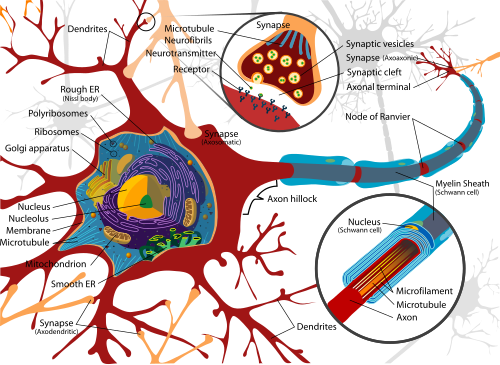
กายวิภาคและวิทยาเนื้อเยื่อ
นิวรอนออกแบบโดยเฉพาะเพื่อประมวลและส่งสัญญาณระหว่างเซลล์ เพราะมีหน้าที่มากมายในส่วนต่าง ๆ ของระบบประสาท เซลล์จึงมีรูปร่าง ขนาด และคุณสมบัติทางเคมีไฟฟ้าต่าง ๆ หลากหลาย ยกตัวอย่างอย่างเช่น
- ตัวเซลล์ (soma) อาจมีขนาดระหว่าง 4-100 ไมโครเมตร เป็นส่วนที่มีนิวเคลียสเหมือนเซลล์ทั่วไป และเป็นส่วนที่สังเคราะห์โปรตีนโดยมาก นิวเคลียสอาจมีขนาดระหว่าง 3-18 ไมโครเมตร
- ใยประสาทนำเข้า/เดนไดรต์เป็นส่วนยื่นออกของเซลล์ที่มีสาขามากมาย รูปลักษณ์และโครงสร้างทั่วไปเรียกโดยอุปลักษณ์ได้ว่า เป็นต้นไม้เดนไดรต์ (คือคล้ายรากไม้) เป็นส่วนที่รับสัญญาณผ่านเดนไดรติก สไปน์เข้าสู่ตัวเซลล์โดยมาก
- แอกซอนเป็นโครงสร้างที่ดูบอบบางกว่าและดูเหมือนกับสายเคเบิลที่ยื่นออกไปเป็นระยะเป็นสิบ ๆ เป็นร้อย ๆ หรืออาจเป็นหมื่น ๆ เท่าของขนาดตัวเซลล์ เป็นตัวส่งสัญญาณประสาทไปจากตัวเซลล์ (แต่ก็นำข้อมูลอะไรบางอย่างกลับไปยังตัวเซลล์ด้วย) นิวรอนจำนวนมากมีแอกซอนแค่อันเดียว แต่ปกติแอกซอนจะแตกสาขามากมาย ทำให้สามารถสื่อสารกับเซลล์เป้าหมายเป็นจำนวนมาก
- จุดที่แอกซอนยื่นออกมาจากตัวเซลล์เรียกว่า axon hillock นอกจากจะเป็นโครงสร้างทางกายวิภาคแล้ว axon hillock ยังเป็นส่วนที่มีช่องโซเดียมเปิดปิดโดยศักย์ไฟฟ้า (voltage-dependent sodium channels) ที่หนาแน่นที่สุด ซึ่งทำให้มันเป็นส่วนที่เร้าง่ายที่สุดในนิวรอนและเป็นโซนที่จุดชนวนการส่งสัญญาณ (spike) ของแอกซอน ถ้ากล่าวถึงคุณสมบัติทางสรีรวิทยาไฟฟ้าของมัน ก็จะกล่าวได้ว่ามันมีขีดเริ่มสร้างศักยะงานที่เป็นขั้วลบที่สุดในเซลล์ แม้ว่า แอกซอนและ axon hillock โดยทั่วไปจะมีหน้าที่เกี่ยวกับการส่งข้อมูล แต่ axon hillock ก็ยังสามารถรับสัญญาณจากนิวรอนอื่น ๆ ได้ด้วย
- ส่วนปลายแอกซอน (axon terminal) จะมีจุดประสานประสาท/ไซแนปส์ (synapse) ซึ่งมีหน้าที่เฉพาะคือปล่อยสารสื่อประสาทออกเพื่อสื่อสารกับเซลล์เป้าหมาย
ส่วนต่าง ๆ ของนิวรอนมองโดยทั่วไปว่า ทำหน้าที่โดยเฉพาะ ๆ แต่ว่า ทั้งเดนไดรต์และแอกซอนก็มักจะทำอะไรที่ต่างไปจากหน้าที่ "หลัก" ของมัน
ทั้งเดนไดรต์และแอกซอนในระบบประสาทกลาง (CNS) โดยทั่วไปจะหนาประมาณ 1 ไมโครเมตร แต่ในระบบประสาทนอกส่วนกลาง (PNS) จะหนากว่า ส่วนตัวเซลล์มีขนาดประมาณ 10-25 ไมโครเมตรและมักจะไม่ใหญ่กว่านิวเคลียสของเซลล์ที่อยู่ในมันมากนัก แอกซอนของเซลล์ประสาทสั่งการของมนุษย์ที่ยาวที่สุดอาจจะยาวกว่า 1 เมตร โดยวิ่งจากกระดูกสันหลังไปยังนิ้วเท้า ส่วนแอกซอนของเซลล์ประสาทรับความรู้สึกอาจวิ่งจากนิ้วเท้าไปถึง posterior column ของไขสันหลัง ซึ่งยาวกว่า 1.5 เมตรในผู้ใหญ่ ส่วนยีราฟอาจมีแอกซอนยาวหลายเมตรวิ่งไปตามคอของมันทั้งหมด
ความรู้เกี่ยวกับหน้าที่ของแอกซอนมาจากการศึกษาแอกซอนยักษ์ของปลาหมึก เพราะศึกษาได้ง่ายที่สุดเพราะมีขนาดยักษ์ (โดยเปรียบเทียบ) คือ หนา 0.5-1 มิลลิเมตร และยาวหลายเซนติเมตร นิวรอนที่พัฒนาเต็มที่แล้วจะอยู่ในสภาวะ postmitotic (G0 phase) อย่างถาวร แต่ว่า งานวิจัยเริ่มต้นตั้งแต่ปี 2545 ก็แสดงว่า นิวรอนสามารถเพิ่มขึ้นทั่วสมองโดยเกิดจากเซลล์ประสาทต้นกำเนิด (neural stem cells) ผ่านกระบวนการกำเนิดเซลล์ (neurogenesis) แม้จะพบเซลล์เช่นนี้ทั่วสมอง แต่ก็ปรากฏมากที่สุดใน subventricular zone และ subgranular zone (ในฮิปโปแคมปัส)


วิทยาเนื้อเยื่อและโครงสร้างภายใน
ก้อนจิ๋ว ๆ จำนวนมากที่เรียกว่า Nissl substance (หรือ Nissl body) จะเห็นได้เมื่อตัวเซลล์ประสาทย้อมด้วยสี (Basophilic dye) โครงสร้างนี้ประกอบด้วยร่างแหเอนโดพลาซึม (endoplasmic reticulum) และไรโบโซมอล อาร์เอ็นเอ (rRNA) ที่สัมพันธ์กัน โดยมีชื่อตามจิตแพทย์และนักประสาทพยาธิวิทยา Franz Nissl (พ.ศ. 2403-2462) และมีหน้าที่เกี่ยวกับการสังเคราะห์โปรตีน ความเด่น/มากมายของมันอธิบายได้ว่า เซลล์ประสาทมีระดับเมแทบอลิซึมสูงมาก จึงต้องสร้างโปรตีนมาก สีชอบภาวะเบสเช่น aniline หรือ haematoxylin อ่อน ๆ จะเน้นโครงสร้างที่มีขั้วลบ โดยเข้ายึดกับโครงฟอสเฟตของ rRNA
ตัวนิวรอนเองมีโครงโปรตีนสนับสนุนที่เรียกว่า neurofilament ซึ่งรวมตัวกันเป็น neurofibril นิวรอนบางอย่างยังมีเม็ดสี เช่น neuromelanin ซึ่งเป็นสีออกน้ำตาล-ดำ และเป็นผลพลอยได้ของการสังเคราะห์ catecholamine, และ lipofuscin ซึ่งเป็นสีออกเหลือง-น้ำตาล โดยเม็ดสีทั้งสองจะเพิ่มขึ้นเรื่อย ๆ ตามอายุ
โปรตีนทางโครงสร้างที่สำคัญต่อหน้าที่ของนิวรอนก็คือ แอกติน และ tubulin ของไมโครทิวบูล แอกติน โดยมากพบที่ปลายแอกซอนและเดนไดรต์เมื่อนิวรอนกำลังพัฒนาขึ้น แต่โครงสร้างภายในของแอกซอนและเดนไดรต์นั้นก็แตกต่างกัน แอกซอนโดยทั่วไปจะไม่มีไรโบโซม ยกเว้นในส่วนต้น ส่วนเดนไดรต์จะมีร่างแหเอนโดพลาซึมหรือไรโบโซมเป็นเม็ด ๆ โดยมีปริมาณน้อยลงไปเรื่อย ๆ เมื่อห่างออกจากตัวเซลล์

การจัดประเภท
นิวรอนมีรูปร่างและขนาดต่าง ๆ กัน ดังนั้นจึงสามารถจัดประเภทตามสัณฐานและหน้าที่ โดยนักกายวิภาค Camillo Golgi จัดกลุ่มนิวรอนไว้ 2 พวก แบบ I มีแอกซอนยาวเพื่อส่งสัญญาณในระยะไกล และแบบ II มีแอกซอนสั้น ซึ่งมักจะสับสนกับเดนไดรต์
แบบ I สามารถจัดย่อยว่าตัวเซลล์อยู่ที่ตำแหน่งไหน สัณฐานพื้นฐานของแบบ I ซึ่งมีตัวอย่างเป็นเซลล์ประสาทสั่งการ (motor neuron) ของไขสันหลัง จะมีตัวเซลล์ (soma) และแอกซอนยาวบางหุ้มปลอกไมอีลิน รอบ ๆ ตัวเซลล์จะเป็นสาขาของต้นไม้เดนไดรต์ที่รับสัญญาณจากนิวรอนอื่น ๆ ส่วนแอกซอนจะไปสุดที่สาขาต่าง ๆ ที่เรียกว่าปลายแอกซอน (axon terminal) ซึ่งปล่อยสารสื่อประสาทเข้าไปในช่องที่เรียกว่าร่องไซแนปส์ (synaptic cleft) ซึ่งอยู่ระหว่างปลายแอกซอนและเดนไดรต์ของเซลล์ต่อไป
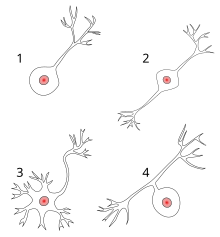
1 นิวรอนขั้วเดียว (Unipolar neuron)
2 นิวรอนสองขั้ว (Bipolar neuron)
3 นิวรอนหลายขั้ว (Multipolar neuron)
4 นิวรอนขั้วเดียวเทียม (Pseudounipolar neuron)
การแบ่งประเภทตามโครงสร้าง
ขั้ว
นิวรอนโดยมากสามารถจัดกลุ่มตามกายวิภาคว่าเป็น
- นิวรอนขั้วเดียว (Unipolar neuron) หรือนิวรอนขั้วเดียวเทียม (Pseudounipolar neuron) มีเดนไดรต์และแอกซอนยื่นออกมาจากขั้วเดียว
- นิวรอนสองขั้ว (Bipolar neuron) มีแอกซอนและเดนไดรต์อันเดียวอยู่ตรงข้ามกันขั้นระหว่างโดยตัวเซลล์
- นิวรอนหลายขั้ว (Multipolar neuron) มีเดนไดรต์ 2 อันหรือมากกว่านั้น และมีแอกซอนต่างหาก
- Golgi I เป็นนิวรอนที่มีแอกซอนยาว ตัวอย่างรวมทั้ง นิวรอนพีระมิด เซลล์เพอร์คินจี และ anterior horn cell
- Golgi II เป็นนิวรอนที่มีแอกซอนส่งไปยังที่ใกล้ ๆ ตัวอย่างที่ดีสุดก็คือ granule cell
- นิวรอนไร้แอกซอน (Anaxonic neuron) มีแอกซอนที่ไม่สามารถจำแนกจากเดนไดรต์
ขนาดแอกซอน
เมื่อแบ่งชนิดตามขนาดแอกซอน จะสามารถแบ่งเซลล์ประสาทออกเป็น 3 ชนิดใหญ่ ๆ ได้แก่ A, B และ C โดยแต่ละชนิดจะจำแนกเป็นกลุ่มย่อยอีก เช่น ชนิด A ก็จะแบ่งเป็นกลุ่ม แอลฟา เบตา แกมมา เดลตา เป็นต้น ซึ่งแต่ละชนิดจะมีลักษณะ คุณสมบัติ และหน้าที่ที่แตกต่างกันออกไป โดยแอกซอนชนิด A จะมีเส้นผ่านศูนย์กลางใหญ่ที่สุด รองลงมาคือชนิด B และ ชนิด C เมื่อวัดความไวการตอบสนองของแอกซอนเหล่านี้ ก็จะพบว่า ที่มีขนาดใหญ่กว่าจะตอบสนองต่อสิ่งเร้าได้เร็วกว่า ดังนั้น เซลล์ชนิด A จึงตอบสนองต่อการกระตุ้นได้เร็วกว่าชนิด B และ C ตามลำดับ สำหรับการตอบสนองต่อภาวะต่าง ๆ ของเซลล์ชนิด A, B และ C จะเป็นดังนี้
- ภาวะขาดออกซิเจน ผลการตอบสนองของ B > A > C
- ภาวะที่มีแรงกด A > B > C
- ภาวะที่มียาชาเฉพาะที่ C > B > A
อื่น ๆ
นอกจากนั้นแล้ว ประเภทของนิวรอนยังแยกได้ตามตำแหน่งในระบบประสาทและรูปร่างของมัน ยกตัวอย่างเช่น
- Basket cell (เซลล์ตะกร้า) เป็นเซลล์ประสาทต่อประสาน (interneuron) ที่มีข่ายแอกซอนหนาแน่นรอบ ๆ ตัวของเซลล์เป้าหมาย พบอยู่ในเปลือกสมองและสมองน้อย
- Betz cell เป็นนิวรอนสั่งการขนาดใหญ่
- Lugaro cell เป็นเซลล์ประสาทต่อประสานของสมองน้อย
- Medium spiny neuron เป็นนิวรอนโดยมากใน corpus striatum
- เซลล์เพอร์คินจี เป็นนิวรอนขนาดใหญ่มากอยู่ในสมองน้อย เป็นนิวรอนหลายขั้วแบบ Golgi I
- เซลล์พิระมิดเป็นนิวรอนที่มีตัวเซลล์เป็นรูปสามเหลี่ยม เป็นนิวรอนหลายขั้วแบบ Golgi I
- Renshaw cell เป็นนิวรอนที่มีขั้วทั้งสองเชื่อมกับเซลล์ประสาทสั่งการแบบอัลฟา
- Unipolar brush cell เป็นเซลล์ประสาทต่อประสานที่มีปลายเดนไดรต์ดูเหมือนกับพุ่มไม้
- Granule cell เป็นนิวรอนหลายขั้วแบบ Golgi II
- Anterior horn cell เป็นเซลล์ประสาทสั่งการ (Motor neuron) ที่อยู่ในไขสันหลัง
- Spindle cell เป็นเซลล์ประสาทต่อประสานที่เชื่อมเขตที่อยู่ห่าง ๆ ในสมอง
ตามหน้าที่
โดยทิศทางของสัญญาณ
- เซลล์ประสาทนำเข้า (Afferent neuron) ส่งข้อมูลจากเนื้อเยื่อและอวัยวะต่าง ๆ ไปยังระบบประสาทกลาง โดยบางครั้งเรียกว่า เซลล์ประสาทรับความรู้สึก (sensory neurons)
- เซลล์ประสาทส่งออก (Efferent neuron) ส่งสัญญาณจากระบบประสาทกลางไปยังเซลล์ปฏิบัติงาน (effector cell) โดยบางครั้งเรียกว่า เซลล์ประสาทสั่งการ (motor neuron)
- เซลล์ประสาทต่อประสาน (interneuron) เชื่อมนิวรอนภายในเขตโดยเฉพาะ ๆ ของระบบประสาทกลาง
แต่ว่า เซลล์ประสาทนำเข้าและเซลล์ประสาทส่งออก ก็ยังหมายถึงนิวรอนที่ทั่วไปยิ่งกว่านั้น ที่นำข้อมูลไปยังหรือส่งข้อมูลไปจากเขตต่าง ๆ ในสมอง
ฤทธิ์ต่อเซลล์ประสาทอื่น ๆ
เซลล์ประสาทออกฤทธิ์ต่อนิวรอนอื่น ๆ โดยปล่อยสารสื่อประสาทที่เข้ายึดกับหน่วยรับความรู้สึกทางเคมี (chemical receptor) แต่ผลที่เกิดกับนิวรอนหลังไซแนปส์ไม่ได้กำหนดโดยนิวรอนก่อนไซแนปส์หรือโดยสารสื่อประสาท แต่โดยประเภทของหน่วยรับความรู้สึกที่ทำงาน สารสื่อประสาทอุปมาเหมือนกับลูกกุญแจ และหน่วยรับความรู้สึกเหมือนแม่กุญแจ ลูกกุญแจแบบเดียวกันสามารถใช้ไขแม่กุญแจหลายแบบ หน่วยรับความรู้สึกสามารถจัดกว้าง ๆ ว่าเป็นแบบเร้า (excitatory) คือเพิ่มอัตราศักยะงาน หรือเป็นแบบยับยั้ง (inhibitory) คือลดอัตราศักยะงาน หรือเป็นแบบควบคุม (modulatory) คือมีผลระยะยาวแต่ไม่มีฤทธิ์โดยตรงกับอัตราการส่งศักยะงาน
สารสื่อประสาทที่สามัญที่สุดในสมองสองอย่าง คือ กลูตาเมตและกรดแกมมาอะมิโนบิวทีริก (กาบา) ออกฤทธิ์โดยมากคล้าย ๆ กัน คือ กลูตาเมตมีฤทธิ์ต่อหน่วยรับความรู้สึกหลายอย่าง โดยมีฤทธิ์เร้าต่อ ionotropic receptor และฤทธิ์ควบคุมต่อ metabotropic receptor และคล้าย ๆ กัน กาบามีฤทธิ์ต่อหน่วยรับความรู้สึกหลายประเภท แต่มีฤทธิ์เหมือนกัน (อย่างน้อยในสัตว์ที่โตแล้ว) คือเป็นตัวยับยั้ง เพราะความสม่ำเสมอเช่นนี้ นักประสาทวิทยาศาสตร์จึงมักใช้คำง่าย ๆ โดยกล่าวถึงเซลล์ที่ปล่อยกลูตาเมตว่าเซลล์ประสาทแบบเร้า (excitatory neuron) และเซลล์ที่ปล่อยกาบาว่าเป็นเซลล์ประสาทแบบยับยั้ง (inhibitory neuron) และเพราะว่านิวรอนเกิน 90% ในสมองไม่ปล่อยกลูตาเมตก็กาบา การจัดประเภทเช่นนี้รวมเอานิวรอนโดยมาก ยังมีนิวรอนประเภทอื่น ๆ ที่มีฤทธิ์สม่ำเสมอต่อเซลล์เป้าหมาย ยกตัวอย่างเช่น เซลล์ประสาทสั่งการแบบเร้า (excitatory motor neuron) ในไขสันหลังที่ปล่อยสาร acetylcholine และเซลล์ประสาทไขสันหลังแบบยับยั้ง (inhibitory spinal neuron) ที่ปล่อยสารไกลซีน
แต่ว่าการแบ่งสารสื่อประสาทว่าเป็นแบบเร้าหรือแบบยับยั้งนั้นจะไม่สมบูรณ์ เพราะมันขึ้นอยู่กับหน่วยรับความรู้สึกทางเคมีของนิวรอนหลังไซแนปส์ โดยหลักแล้ว นิวรอนตัวเดียวกันที่ปล่อยสารสื่อประสาทอย่างเดียว สามารถมีฤทธิ์เร้าต่อเป้าหมายบางอย่าง มีฤทธิ์ยับยั้งต่อเป้าหมายบางอย่าง และมีฤทธิ์ควบคุมต่อเป้าหมายที่เหลือ
ยกตัวอย่างเช่น เซลล์รับแสง (photoreceptor cell) ในจอตาจะปล่อยสารสื่อประสาทกลูตาเมตออกเรื่อย ๆ ถ้าไม่มีแสง และจะมีฤทธิ์เร้าต่อเซลล์ประสาทที่เป็นเป้าหมาย คือ OFF Retinal bipolar cell โดยคล้ายกับนิวรอนอื่น ๆ แต่ว่า กลับมีฤทธิ์ยับยั้งต่อเซลล์เป้าหมายที่อยู่ข้าง ๆ ที่เรียกว่า ON Retinal bipolar cell เพราะว่า เซลล์เป้าหมายไม่มี ionotropic glutamate receptor และกลับมี inhibitory metabotropic glutamate receptor เมื่อมีแสง เซลล์รับแสงก็จะหยุดปล่อยกลูตาเมต ซึ่งระงับการยับยั้งของ ON bipolar cell และทำให้มันเริ่มส่งศักยะงาน แต่นี่ก็จะระงับการเร้า OFF bipolar cell ไปพร้อม ๆ กัน และทำให้มันหยุดส่งศักยะงาน
ถึงกระนั้น ก็ยังสามารถระบุผลยับยั้งของนิวรอนก่อนไซแนปส์ที่จะมีต่อนิวรอนหลังไซแนปส์ได้ โดยขึ้นกับโปรตีนที่แสดงออกโดยนิวรอนก่อนไซแนปส์ นิวรอนที่แสดงออก parvalbumin ปกติจะลดการส่งสัญญาณของนิวรอนหลังไซแนปส์ในเปลือกสมองส่วนการเห็น เทียบกับนิวรอนที่แสดงออก somatostatin ซึ่งปกติจะระงับการรับข้อมูลที่เดนไดรต์ของนิวรอนหลังไซแนปส์
รูปแบบการส่งสัญญาณ
นิวรอนยังสามารถจัดตามคุณสมบัติทางสรีรวิทยาไฟฟ้า คือ
- ส่งสัญญาณเรื่อย ๆ (Tonic or regular spiking) นิวรอนบางอย่างปกติส่งสัญญาณเรื่อย ๆ ยกตัวอย่างเช่น เซลล์ประสาทต่อประสาน (interneuron) ของ neurostriatum
- ส่งสัญญาณเป็นชุด ๆ (Phasic or bursting) นิวรอนที่ส่งสัญญาณเป็นชุด ๆ เรียกว่า phasic neuron
- ส่งสัญญาณเร็ว (Fast spiking) นิวรอนบางอย่างเด่นตรงที่อัตราการส่งสัญญาณสูง ยกตัวอย่างเช่น เซลล์ประสาทต่อประสาทแบบยับยั้งในเปลือกสมองบางอย่าง เซลล์ใน globus pallidus และ Retinal ganglion cell
โดยการผลิตสารสื่อประสาท
- Cholinergic neuron ผลิตสารสื่อประสาท acetylcholine โดยปล่อยออกจากนิวรอนก่อนไซแนปส์เข้าไปในร่องไซแนปส์ (synaptic cleft) และสารทำงานเป็นลิแกนด์ที่เข้ายึดกับช่องไอออนเปิดปิดโดยลิแกนด์ (ligand-gated ion) และ metabotropic (GPCRs) muscarinic receptor เช่นหน่วยรับนิโคติน (Nicotinic receptor) เป็นช่องไอออนเปิดปิดโดยลิแกนด์แบบมีหน่วยโปรตีนย่อย 5 หน่วย (pentameric) โดยหน่วยย่อยมีทั้งแบบอัลฟาและบีตาที่ยึดกับนิโคติน การเข้ายึดของลิแกนด์จะเปิดช่องไอออนทำให้เกิดการลดขั้ว (depolarization) เนื่องจากไอออน Na+ ไหลเข้าไปในเซลล์ และเพิ่มโอกาสที่เซลล์ก่อนไซแนปส์จะปล่อยสารสื่อประสาท สาร Acetylcholine สังเคราะห์มาจาก choline และอะซิติลโคเอนไซม์ เอ
- GABAergic neuron ผลิตกรดแกมมาอะมิโนบิวทีริก (กาบา) กาบาเป็นสารสื่อประสาทแบบยับยั้งอย่างหนึ่งในสองอย่างในระบบประสาทกลาง อีกอย่างหนึ่งคือไกลซีน กาบามีหน้าที่เหมือนกับ ACh คือเปิดช่องแอนไอออน (anion channel) ที่ปล่อยให้ไอออน Cl− ไหลเข้าไปในเซลล์หลังไซแนปส์ Cl− เป็นเหตุให้เกิดสภาวะ hyperpolarization ภายในนิวรอน ลดโอกาสที่เซลล์จะยิงศักยะงานเพราะศักย์ไฟฟ้ามีค่าลบมากยิ่งขึ้น (คือถ้าจะมีการยิงศักยะงาน ขีดเริ่มเปลี่ยนศักย์ไฟฟ้าจะต้องมีค่าบวก) กาบาสังเคราะห์มาจากสารสื่อประสาทกลูตาเมตผ่านเอนไซม์ glutamate decarboxylase
- Glutamatergic neuron ผลิตสารสื่อประสาทกลูตาเมต กลูตาเมตเป็นสารสื่อประสาทที่เป็นกรดอะมิโนแบบเร้าหลักอย่างหนึ่งในสองอย่าง อีกอย่างก็คือ aspartate มีหน่วยรับกลูตามเมต (glutamate receptor) 4 ประเภท ประเภทหนึ่งเป็น G-protein coupled receptor (มักเรียกว่า GPCR) และอีก 3 ประเภทเป็นช่องไอออนเปิดปิดด้วยลิแกนด์ คือ
-
- AMPA receptor และ Kainate receptor ทั้งสองทำหน้าที่เป็นช่องแคตไอออนที่เปิดรับ Na+ อำนวยให้มีการสื่อประสาทผ่านไซแนปส์แบบเร้าอย่างรวดเร็ว
- NMDA receptor เป็นช่องแคนไออนอีกอย่างแต่เปิดรับ Ca2+ ได้ดีกว่า หน้าที่ของ NMDA receptor ขึ้นอยู่กับหน่วยรับไกลซีนซึ่งทำงานร่วมกัน (co-agonist) ดังนั้น NMDA receptor จะทำงานก็ต่อเมื่อมีลิแกนต์ทั้งสองแบบ
- Metabotropic receptor (GPCR) เป็นตัวควบคุม/ปรับการส่งสัญญาณและความเร้าได้ของเซลล์หลังไซแนปส์
- ถ้าขาดเลือดไปเลี้ยงสมอง กลูตาเมตสามารถเร้าเซลล์จนเป็นพิษ (excitotoxicity) มีผลทำให้สมองเสียหาย คือ เมื่อขาดเลือด นิวรอนก่อนไซแนปส์จะปล่อยกลูตาเมตซึ่งจะเร้าหน่วยรับ NMDA และ AMPA มากกว่าปกติ ทำให้ Ca2+ และ Na+ เข้าไปในนิวรอนหลังไซแนปส์ในระดับที่สูงขึ้นแล้วทำให้เซลล์เสียหาย กลูตาเมตสังเคราะห์มาจากกรดอะมิโนกลูตามีนผ่านเอนไซม์ Glutamine oxoglutarate aminotransferase (หรือเรียกว่า glutamate synthase)
- Dopaminergic neuron ผลิตสารโดพามีน ซึ่งเป็นสารสื่อประสาทที่มีฤทธิ์ต่อ GPCR ประเภท D1 (รวมทั้ง D1 และ D5) ซึ่งเพิ่ม cAMP (Cyclic adenosine monophosphate) และ PKA (Protein kinase A), และประเภท D2 (รวม D2, D3, และ D4) ซึ่งมีฤทธิ์ต่อ GPCR ที่ลด cAMP และ PKA โดพามีนสัมพันธ์กับพื้นอารมณ์และพฤติกรรม และยังควบคุมการสื่อประสาทของทั้งเซลล์ก่อนไซแนปส์และหลังไซแนปส์ การสูญเสียนิวรอนแบบโดพามีนในเขต substantia nigra สัมพันธ์กับโรคพาร์คินสัน โดพามีนสังเคราะห์มาจากกรดอะมิโนไทโรซีน คือเอนไซม์ tyrosine hydroxlase จะเร่งปฏิกิริยาเปลี่ยนไทโรซีนไปเป็น levadopa และกรดอะมิโน decarboxylase จะเปลี่ยน levadopa ไปเป็นโดพามีน
- Serotonergic neuron ผลิตสารเซโรโทนิน และเซโรโทนิน (5-Hydroxytryptamine, 5-HT) สามารถมีฤทธิ์เร้าหรือยับยั้ง ในบรรดาหน่วยรับประเภท 5-HT 4 อย่าง 3 อย่างเป็น GPCR และอีกอย่างหนึ่งเป็นช่องแคตไอออนเปิดปิดโดยลิแกนต์ (ligand gated cation channel) เซโรโทนินสังเคราะห์จากทริปโตเฟนผ่านเอนไซม์ tryptophan hydroxylase และต่อมากรดแอโรเมติก decarboxylase การขาดเซโรโทนนินในเซลล์ประสาทหลังไซแนปส์สัมพันธ์กับโรคซึมเศร้า ดังนั้น ยาเช่น ฟลูอ๊อกซิติน และ Zoloft ที่ยับยั้งการนำเซโรโทนินไปใช้ใหม่โดยยับยั้งตัวขนส่งเซโรโทนิน (serotonin transporter) จึงสามารถใช้รักษาโรคได้

หน้าที่และการทำงานของเซลล์ประสาทโดยย่อ
- เซลล์จะมีการสร้างหรือเปลี่ยนแปลงศักย์ไฟฟ้าบนตัวเซลล์อยู่ตลอดเวลา
- รับกระแสประสาทจากเซลล์ประสาทเซลล์อื่นผ่านเดนไดรต์
- ประมวลสัญญาณประสาทเข้า ซึ่งถ้าแอมพลิจูดของศักย์ไฟฟ้าสูงถึงจุด (เยอรมัน: Schwellenpotential อังกฤษ: threshold potential) ก็จะชักนำให้เกิดศักยะงานขึ้น (เยอรมัน: Aktionspotential อังกฤษ: action potential)
- แปลรหัสให้เกิดกระแสประสาทที่บริเวณ axon hillock ซึ่งกระแสประสาทก็จะแล่นไปตามแอกซอน โดยความถี่ของกระแสประสาทนั้นจะเกิดเป็นชุด ๆ ตามลักษณะของเซลล์ (ดู รูปแบบการส่งสัญญาณ)
- ส่งต่อข้อมูลสัญญาณประสาทให้กับเซลล์ประสาทเซลล์อื่นต่อไปผ่านจุดประสานประสาท
การเชื่อมต่อ
นิวรอนสื่อสารกันเองผ่านจุดประสานประสาท (ไซแนปส์) โดยปลายแอกซอน (axon terminal หรือ en passant bouton) ของเซลล์หนึ่งจะอยู่ติดกับเดนไดรต์ ตัวเซลล์ หรือ (แม้จะน้อยกว่า) แอกซอนของอีกเซลล์หนึ่ง นิวรอนเช่นเซลล์เพอร์คินจีในสมองน้อยอาจมีสาขาเดนไดรต์เป็นพัน โดยเชื่อมกับเซลล์อื่น ๆ อีกเป็นหมื่น ๆ นิวรอนอื่น ๆ เช่น magnocellular neuron ของ supraoptic nucleus (ในไฮโปทาลามัส) มีเดนไดรต์เพียแค่ 1–2 สาขา แต่ว่าแต่ละสาขามีไซแนปส์เป็นพัน ๆ ไซแนปส์สามารถเป็นทั้งแบบเร้า (excitatory, EPSP) หรือแบบยับยั้ง (inhibitory, IPSP) และสามารถเพิ่มหรือลดการทำงานของเซลล์เป้าหมายตามลำดับ มีนิวรอนที่สามารถสื่อสารผ่านจุดประสานประสาทไฟฟ้า ผ่านช่อง gap junction ที่สามารถนำไฟฟ้าระหว่างเซลล์
ในไซแนปส์เคมี กระบวนการสื่อประสาทเป็นดังต่อไปนี้ เมื่อศักยะงานวิ่งมาถึงปลายแอกซอน (axon terminal) เซลล์ก็จะเปิดช่องแคลเซียมที่เปิดปิดโดยศักย์ไฟฟ้า (voltage-gated calcium channel) ทำให้ไอออนแคลเซียมไหลเข้ามาในปลายแอกซอนได้ แคลเซียมจะทำให้ถุงไซแนปส์ (synaptic vesicle) จำนวนหนึ่งที่เต็มไปด้วยโมเลกุลสารสื่อประสาท เชื่อมเข้ากับเยื่อหุ้มเซลล์ แล้วปล่อยสารเขาไปในร่องไซแนปส์ (synaptic cleft) สารก็จะแพร่ข้ามร่องไซแนปส์และออกฤทธิ์กับตัวรับของนิวรอนหลังไซแนปส์ นอกจากนั้นแล้ว ระดับแคลเซียมที่สูงขึ้นในไซโทพลาซึมที่ปลายแอกซอน (axon terminal) ยังจุดชนวนให้ไมโทคอนเดรียดูดซึมแคลเซียม (mitochondrial calcium uptake) ซึ่งก็จะเริ่มกระบวนการเมแทบอลิซึมทางพลังงานของไมโทคอนเดรียเพื่อผลิดอะดีโนซีนไตรฟอสเฟต (ATP) เพื่อเป็นพลังงานดำรงการสื่อประสาท
สมองมนุษย์มีไซแนปส์จำนวนมหาศาล นิวรอนแต่ละตัวในแสนล้านตัว (1011) มีไซแนปส์ 7,000 อันที่เชื่อมกับนิวรอนอื่น ๆ โดยเฉลี่ย สมองของเด็กอายุสามขวบประเมินว่า มีไซแนปส์ประมาณพันล้านล้านอัน (1015) แต่จำนวนจะลดลงตามอายุ แล้วคงจำนวนเมื่อถึงวัยผู้ใหญ่ ค่าประเมินในผู้ใหญ่อยู่ระหว่าง 100-500 ล้านล้านไซแนปส์ (1014-5 x 1014)

กลไกในการส่งต่อศักยะงาน
ในปี 2480 นักประสาทสรีรวิทยาชาวอังกฤษ ดร. จอห์น แซคารี่ ยัง เสนอว่าแอกซอนยักษ์ของปลาหมึกสามารถใช้ศึกษาคุณสมบัติทางไฟฟ้าของเซลล์ประสาทได้ เพราะแม้ว่าจะใหญ่กว่า แต่ก็ยังมีคุณสมบัติคล้ายกับนิวรอนมนุษย์ ดังนั้น จึงศึกษาได้ง่ายกว่า โดยใส่อิเล็กโทรดเข้าไปในแอกซอนยักษ์ ก็จะสามารถวัดศักย์เยื่อหุ้มเซลล์ได้อย่างแม่นยำ
เยื่อหุ้มเซลล์ทั้งที่แอกซอนและตัวเซลล์ มีช่องไอออนเปิดปิดโดยศักย์ไฟฟ้า (voltage-gated ion channel) ที่ทำให้นิวรอนสามารถสร้างและส่งสัญญาณทางไฟฟ้าโดยเป็นศักยะงาน ซึ่งสร้างขึ้นและส่งต่อโดยใช้ไอออนที่มีประจุไฟฟ้ารวมทั้งโซเดียม (Na+) โพแทสเซียม (K+) คลอไรด์ (Cl−) และแคลเซียม (Ca2+)
มีสิ่งเร้าหลายอย่างที่สามารถทำให้นิวรอนเกิดการทำงานทางไฟฟ้า รวมทั้งสัมผัส/ความดัน ความยืด สารเคมี และความเปลี่ยนแปลงต่อความต่างศักย์ข้ามเยื่อหุ้มเซลล์ สิ่งเร้าจะเป็นเหตุให้ช่องไอออนโดยเฉพาะที่ฝังอยู่ในเยื่อหุ้มเซลล์เปิด ทำให้มีไอออนไหลผ่านเยื่อ แล้วเปลี่ยนความต่างศักย์ของเยื่อหุ้มเซลล์
แม้ว่า นิวรอนและแอกซอนที่บาง ๆ จะมีระดับเมแทบอลิซึมที่น้อยกว่า (ใช้พลังงานน้อยกว่า) เพื่อสร้างและส่งศักยะงาน แต่ว่า แอกซอนที่หนากว่าสามารถส่งพัลส์ไฟฟ้าได้เร็วกว่า เพื่อลดระดับเมแทบอลิซึมให้มากที่สุดและส่งสัญญาณให้เร็วที่สุด นิวรอนจำนวนมากมีปลอกไมอีลินหุ้มแอกซอน ซึ่งเกิดจากเซลล์เกลีย คือโอลิโกเดนโดรไซต์ในระบบประสาทกลาง (CNS) และเซลล์ชวานน์ในระบบประสาทนอกส่วนกลาง (PNS) ปลอกช่วยให้ศักยะงานวิ่งไปได้เร็วกว่าแม้ในแอกซอนที่มีขนาดเดียวกัน และยังใช้พลังงานน้อยกว่าอีกด้วย ปลอกไมอีลินใน PNS มักจะแล่นไปตามแอกซอนโดยแบ่งออกเป็นส่วน ๆ แต่ละส่วนยาวประมาณ 1 มม. ขั้นโดย node of Ranvier ที่ไม่ได้หุ้มปลอก แต่มีช่องไอออนเปิดปิดโดยศักย์ไฟฟ้าอย่างหนาแน่น โรคปลอกประสาทเสื่อมแข็ง (MS) เป็นโรคทางประสาทที่เกิดจากการลอกปลอกไมอีลินของแอกซอนใน CNS
แต่ว่า ก็ยังมีนิวรอนที่ไม่สร้างศักยะงานเพื่อส่งสัญญาณ แต่สร้าง Graded potential (ศักย์มีหลายระดับ) ซึ่งก็จะทำให้เซลล์ปล่อยสารสื่อประสาทในหลายระดับเช่นกัน นิวรอนที่ไม่ส่งสัญญาณแบบมียอดแหลม (nonspiking neuron) เช่นนี้ มักจะเป็นเซลล์ประสาทรับความรู้สึกและเซลล์ประสาทต่อประสาน (interneuron) เพราะว่าไม่สามารถส่งสัญญาณไปได้ไกล ๆ
รหัสการส่งสัญญาณ
การเข้ารหัสประสาท (Neural coding) เป็นศาสตร์ที่สนใจว่า นิวรอนเข้ารหัสข้อมูลทางประสาทสัมผัสและข้อมูลอื่น ๆ เพื่อแสดงกับสมองเช่นไร โดยมีจุดหมายหลักเพื่อกำหนดความสัมพันธ์ระหว่างสิ่งเร้ากับการตอบสนองของนิวรอนหรือกลุ่มนิวรอน (neural ensemble) และความสัมพันธ์ของการทำงานทางไฟฟ้าของนิวรอนภายในกลุ่ม เชื่อว่า นิวรอนสามารถเข้ารหัสทั้งข้อมูลดิจิทัลและสัญญาณแอนะล็อก

หลักมีหรือไม่มี
กระแสประสาทเป็นตัวอย่างของการตอบสนองแบบมีหรือไม่มี (all-or-none) คือ ถ้านิวรอนตอบสนอง ก็จะตอบสนองอย่างบริบูรณ์ การเร้าในระดับที่สูงกว่าจะไม่ทำให้เกิดสัญญาณที่แรงกว่า แต่อาจทำให้เกิดสัญญาณที่ส่งถี่กว่า
หน่วยรับความรู้สึก (receptor) มีการตอบสนองหลายอย่างต่อสิ่งเร้า ไม่ว่าจะเป็นแบบปรับตัวอย่างช้า ๆ หรือส่งสัญญาณเรื่อย ๆ (tonic) ที่ตอบสนองต่อสิ่งเร้าที่สม่ำเสมอโดยส่งสัญญาณในอัตราที่สม่ำเสมอ แบบส่งสัญญาณเรื่อย ๆ มักจะตอบสนองต่อสิ่งเร้าที่แรงขึ้นโดยเพิ่มความถี่การยิงสัญญาณ ปกติเป็นกราฟฟังก์ชันยกกำลังของสิ่งเร้าวาดคู่กับอัตราอิมพัลส์ต่อวินาที นี่อุปมาเหมือนกับคุณสมบัติของแสง (ที่ความถี่ใดความถี่หนึ่งโดยเฉพาะ) ที่เมื่อแสงจ้าขึ้น ก็จะต้องมีโฟตอนมากขึ้น เพราะว่า โฟตอนเองไม่สามารถจะ "แรงขึ้น" ที่ความถี่นั้น ๆ
มีตัวรับความรู้สึกบางชนิดที่ปรับตัวอย่างรวดเร็วที่บางครั้งเรียกว่า phasic receptor ที่การส่งสัญญาณจะลดลงหรือหยุดถ้าสิ่งเร้าคงสม่ำเสมอ ยกตัวอย่างเช่น ผิวหนังมนุษย์เมื่อกระทบกับวัตถุจะทำให้นิวรอนยิงสัญญาณ แต่ถ้าวัตถุมีความดันที่สม่ำเสมอกับผิวหนัง นิวรอนก็จะหยุดยิงสัญญาณ นิวรอนของผิวหนังและกล้ามเนื้อที่ตอบสนองต่อความกดดันและความสั่นจะมีโครงสร้างที่เป็นตัวช่วยกรองข้อมูล
pacinian corpuscle (ดูรูป) เป็นตัวอย่างโครงสร้างแบบนี้ มันมีชั้นรูปกลม ๆ คล้ายกับหอม ซึ่งเกิดขึ้นรอบ ๆ ปลายแอกซอน (axon terminal) เมื่อมีแรงกดดันซึ่งทำให้ตัว corpuscle แปรรูปไป สิ่งเร้าที่เป็นแรงกลจะส่งผ่านไปยังแอกซอนซึ่งจะยิงสัญญาณไฟฟ้า แต่ถ้าแรงกดสม่ำเสมอ ก็จะไม่มีการเร้าอีกต่อไป ดังนั้น โดยทั่วไปแล้ว นิวรอนเช่นนี้ตอบสนองด้วยการลดขั้วชั่วคราวในระยะแรกของการแปรรูป และอีกครั้งหนึ่งเมื่อแรงกดดันหายไปซึ่งเป็นเหตุให้เปลี่ยนรูปอีกครั้งหนึ่ง ยังมีการปรับตัวสำคัญแบบอื่น ๆ อีกที่เพิ่มสมรรถภาพของนิวรอนต่าง ๆ

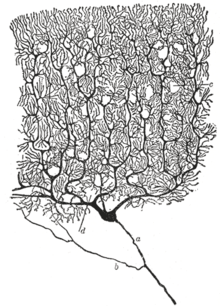
ประวัติ
คำว่า นิวรอน บัญญัติโดยนักกายวิภาคชาวเยอรมัน Heinrich Wilhelm Gottfried von Waldeyer-Hartz โดยมาจากคำในภาษากรีก νεῦρον (neûron) ที่แปลว่า "เส้นด้าย" ส่วนความสำคัญของมันว่าเป็นหน่วยพื้นฐานของระบบประสาทเกิดขึ้นในช่วงต้นคริสต์ทศวรรษที่ 20 ผ่านงานของนักกายวิภาคชาวสเปน นพ. ซานเตียโก รามอน อี กาฆัล ผู้เสนอว่า นิวรอนเป็นเซลล์ต่างหาก ๆ ที่สื่อสารกับกันและกันผ่านจุดประสาน/ช่องว่างที่ออกแบบโดยเฉพาะ หลักนี้ต่อมาเรียกว่า neuron doctrine (หลักนิวรอน) ซึ่งเป็นทฤษฎีหลักทฤษฎีหนึ่งของประสาทวิทยาศาสตร์ปัจจุบัน
เพื่อที่จะดูโครงสร้างของนิวรอนแต่ละตัว ๆ นพ. รามอน อี กาฆัลได้ปรับปรุงกระบวนการย้อมสีด้วยเงินที่รู้จักกันว่า Golgi's method ซึ่งจริง ๆ พัฒนาโดยคู่แข่งของเขาคือ นพ. Camillo Golgi การปรับปรุงของ นพ. รามอน อี กาฆัล ซึ่งรวมเทคนิคที่เขาเรียกว่า double impregnation (การอัดสีสองรอบ) ก็ยังคงใช้อยู่ในปัจจุบัน วิธีการย้อมสีด้วยเงินเป็นวิธีที่มีประโยชน์มากในการตรวจสอบกายวิภาคของระบบประสาท เพราะว่า มันย้อมสีเซลล์ในอัตราที่ต่ำ (แม้ว่าจะไม่รู้ว่าเหตุไร) ดังนั้น จึงสามารถเห็นโครงสร้างเล็ก ๆ ของนิวรอนแต่ละตัวอย่างสมบูรณ์โดยที่ไม่คาบเกี่ยวกับเซลล์ข้าง ๆ มากมายที่อยู่ในเนื้อเยื่อสมองอัดเต็มไปด้วยเซลล์
หลักนิวรอน
หลักนิวรอน (neuron doctrine) เป็นไอเดียพื้นฐานว่า นิวรอนเป็นหน่วยโครงสร้างและหน่วยการทำงานพื้นฐานของระบบประสาท ซึ่งเสนอโดย นพ. ซานเตียโก รามอน อี กาฆัล ในปลายคริสต์ศตวรรษที่ 19 และอ้างว่า นิวรอนเป็นเซลล์โดยเฉพาะ ๆ (ที่ไม่ได้เชื่อมกันเป็นตาข่าย) โดยทำงานแยกส่วนกันทางเมแทบอลิซึม
การค้นพบต่อ ๆ มาจึงเพิ่มรายละเอียดให้กับหลัก ยกตัวอย่างเช่น เซลล์เกลียแม้จะไม่ใช่นิวรอน ก็ยังมีบทบาทสำคัญในการประมวลข้อมูล นอกจากนั้นแล้ว ไซแนปส์ไฟฟ้ายังสามัญกว่าที่เคยคิด คือ นิวรอนเช่นนี้เชื่อมกันโดยตรงและแชร์ไซโทพลาซึมร่วมกัน และจริง ๆ แล้วก็มีตัวอย่างของนิวรอนที่ทำงานร่วมกันอย่างสนิทยิ่งกว่านั้น เช่น แอกซอนยักษ์ของปลาหมึกเกิดจากการรวมกันของแอกซอนหลายอัน
รามอน อี กาฆัล ยังตั้งกฎ Law of Dynamic Polarization ซึ่งอ้างว่า นิวรอนรับสัญญาณมาจากเดนไดรต์และตัวเซลล์ และส่งมันเป็นศักยะงานไปตามแอกซอนโดยทางเดียว คือไปจากตัวเซลล์ โดยมีข้อยกเว้นที่สำคัญ เพราะว่า เดนไดรต์ก็สามารถส่งข้อมูลที่ไซแนปส์ได้เหมือนกัน และแอกซอนก็สามารถรับข้อมูลที่ไซแนปส์
นิวรอนในสมอง
จำนวนนิวรอนในสมองต่างกันอย่างสำคัญในสปีชีส์ต่าง ๆสมองมนุษย์มีนิวรอนประมาณ 85,000-86,000 ล้านตัว โดยมี 16,300 ล้านตัวในเปลือกสมองและ 69,000 ล้านตัวในสมองน้อย โดยเทียบกันแล้ว หนอนนีมาโทดา Caenorhabditis elegans มีนิวรอนเพียงแค่ 302 ตัว ทำให้มันเป็นสัตว์ทดลองที่ดีมาก และนักวิทยาศาสตร์ได้สร้างแผนที่ของนิวรอนของหนอนทั้งหมดแล้ว ส่วนสัตว์ทดลองทางชีววิทยาที่สามัญอีกอย่างหนึ่ง คือ แมลงวันทอง (Drosophila melanogaster) มีนิวรอนประมาณแสนตัว และมีพฤติกรรมซับซ้อนมากมาย
คุณสมบัติหลายอย่างของนิวรอน เริ่มตั้งแต่สารสื่อประสาทและช่องไอออนที่ใช้ เหมือนกันในสปีชีส์ต่าง ๆ ทำให้นักวิทยาศาสตร์สามารถศึกษากระบวนการที่เกิดในสิ่งมีชีวิตที่ซับซ้อนในระบบการทดลองที่ซับซ้อนน้อยกว่า
โรคในระบบประสาท
Charcot-Marie-Tooth disease (CMT) หรือรู้จักว่า hereditary motor and sensory neuropathy (HMSN) หรือ hereditary sensorimotor neuropathy หรือ peroneal muscular atrophy เป็นโรคทางพันธุกรรมหลายชนิดที่มีผลต่อระบบประสาท กำหนดโดยการเสียกล้ามเนื้อและความรู้สึกสัมผัส โดยหลักที่เท้าและขา แต่ก็เกิดที่มือและแขนในระยะหลัง ๆ ด้วย เป็นโรคที่ปัจจุบันยังรักษาไม่ได้ และเป็นโรคทางประสาทสืบทอดทางกรรมพันธุ์ที่สามัญที่สุด มีผลต่อประชากรในอัตรา 37 / 100,000 คน
โรคอัลไซเมอร์ (AD) เป็นโรคประสาทเสื่อม (neurodegenerative disease) กำหนดโดยความเสื่อมทางการรู้คิดอย่างต่อเนื่อง มีสมรรถภาพในการใช้ชีวิตที่ลดลง และมีอาการทางประสาทและพฤติกรรมอื่น ๆ อาการเบื้องต้นที่เด่นที่สุดก็คือการเสียความจำระยะสั้น (ภาวะเสียความจำ) ซึ่งปกติปรากฏเป็นการหลงลืมอะไรเล็ก ๆ น้อย ๆ แล้วแย่ลง แม้ว่าจะไม่เสียความจำเก่า ๆ เมื่อโรคแย่ลง ความพิการทางการรู้คิดจะขยายไปในด้านภาษา (ภาวะเสียการสื่อความ [aphasia]) การเคลื่อนไหวที่ละเอียด (ภาวะเสียการรู้ปฏิบัติ [apraxia]) การรู้จำ (ภาวะเสียการระลึกรู้ [agnosia]) และสมรรถภาพอื่น ๆ เช่นการตัดสินใจและการวางแผน
โรคพาร์คินสัน (PD) เป็นโรคประสาทเสื่อมในระบบประสาทกลาง (CNS) ที่บ่อยครั้งทำให้การเคลื่อนไหวและการพูดพิการ เป็นโรคในกลุ่มที่เรียกว่าโรคทางการเคลื่อนไหว (movement disorders) และกำหนดโดยสภาพแข็งแกร็งของกล้ามเนื้อ การสั่น การเคลื่อนไหวที่ช้าลง (bradykinesia) และในกรณีที่รุนแรง การขยับไม่ได้ (akinesia) อาการปฐมภูมิเป็นผลจากการที่เขต basal ganglia เร้าเปลือกสมองเขตสั่งการ (motor cortex) น้อยลง ปกติเพราะมีสารโดพามีนน้อยหรือไม่ทำงาน โดย dopaminergic neuron เป็นเซลล์ประสาทที่ผลิตสาร อาการทุติยภูมิอาจเป็นความพิการทางการรู้คิดในระดับสูงและปัญหาทางภาษาที่ละเอียดอื่น ๆ PD เป็นโรคทั้งเรื้อรังและลุกลาม
โรคกล้ามเนื้ออ่อนแรงชนิดร้าย (MG) เป็นโรคประสาทกล้ามเนื้อที่ทำให้กล้ามเนื้ออ่อนเปลี้ยและเหนื่อยง่ายเป็นระยะ ๆ แม้ทำกิจกรรมแบบเบา ๆ มีเหตุจากสารภูมิต้านทาน (antibodies) ที่เข้าไปขวาง acetylcholine receptor และที่ต่อประสาทกล้ามเนื้อ (neuromuscular junction) หลังไซแนปส์ และยับยั้งฤทธิ์เร้าของสารสื่อประสาท MG รักษาได้ด้วยยากดภูมิต้านทาน (immunosuppressant), cholinesterase inhibitor, และในบางกรณีการตัดต่อมไทมัส (thymectomy)

การเสียปลอกไมอีลิน
Demyelination เป็นการเสียปลอกไมอีลินที่เป็นฉนวนสำหรับใยประสาท เมื่อปลอกไมอีลินเสื่อมลง สัญญาณที่ส่งไปตามเส้นประสาทอาจเสียหายหรือสูญไป และในที่สุดเส้นประสาทก็จะฝ่อ ซึ่งนำไปสู่โรคประสาทเสื่อมบางอย่าง เช่น โรคปลอกประสาทเสื่อมแข็ง และ chronic inflammatory demyelinating polyneuropathy
แอกซอนเสื่อม
แม้ว่า การตอบสนองต่อความบาดเจ็บโดยมากจะรวมการเพิ่มระดับแคลเซียมเพื่ออุดส่วนที่แตก/ขาด การบาดเจ็บต่อแอกซอนเบื้องต้นจะทำให้เกิดการเสื่อมแบบฉับพลัน คือการขาดออกจากกันระหว่างต้นและปลายของแอกซอนภายใน 30 นาทีของการบาดเจ็บ การเสื่อมจะตามมาด้วยการบวมของเยื่อหุ้มแอกซอน (axolemma) ซึ่งต่อมาจะกลายเป็นเม็ด ๆ การแตกสลายเป็นเม็ด ๆ ของระบบเส้นใยของเซลล์ (cytoskeleton) และออร์แกเนลล์จะเกิดหลังเยื่อหุ้มแอกซอนเสื่อม ความเปลี่ยนแปลงในเบื้องต้นรวมทั้งการคั่งของของไมโทคอนเดรียที่เขต paranodal region ณ ที่บาดเจ็บ ร่างแหเอนโดพลาซึม (ER) จะเสื่อมและไมโทคอนเดรียจะบวมขึ้นแล้วก็แตกสลายไป
ความเสื่อมจะขึ้นอยู่กับโปรตีน ubiquitin และ calpain proteases (ซึ่งมีเหตุมาจากการไหลเข้าของไอออนแคลเซียม) ซึ่งแสดงว่า การเสื่อมของแอกซอนเป็นกระบวนการที่ต้องอาศัยเมแทบอลิซึม ด้วยกระบวนการนี้ แอกซอนจะสลายออกเป็นส่วน ๆ ซึ่งมักจะใช้เวลาประมาณ 24 ชม. ใน PNS และนานกว่านั้นใน CNS แต่ขั้นตอน signaling pathways ที่นำไปสู่การสลายของเยื่อหุ้มแอกซอน (axolemma) ปัจจุบันยังไม่ชัดเจน
การคืนสภาพและการเกิดของประสาท
มีหลักฐานแล้วว่ากำเนิดของประสาท (neurogenesis) สามารถเกิดขึ้นได้ในสมองสัตว์มีกระดูกสันหลังที่โตแล้ว ซึ่งเป็นเรื่องที่สร้างความขัดแย้งในปี 2542 งานศึกษาเกี่ยวกับอายุของนิวรอนมนุษย์ต่อ ๆ มาแสดงว่า กระบวนการนี้เกิดขึ้นกับเซลล์ส่วนน้อย และนิวรอนส่วนมากที่อยู่ใน neocortex จะสร้างขึ้นก่อนคลอดแล้วคงสภาพอยู่โดยไม่มีการเปลี่ยน บ่อยครั้ง แอกซอนส่วนปลายสามารถงอกใหม่ได้ถ้าขาด
งานศึกษาเร็ว ๆ นี้แสดงว่า ร่างกายมีเซลล์ต้นกำเนิดหลายชนิดที่มีสมรรถภาพเพื่อพัฒนาเป็นนิวรอน รายงานในวารสาร Nature ปี 2554 แสดงว่า นักวิจัยพบวิธีการเปลี่ยนเซลล์ผิวหนังมนุษย์ให้เป็นเซลล์ประสาทที่ใช้งานได้ ผ่านกระบวนการ transdifferentiation ที่ "บังคับให้เซลล์รับเอาเอกลักษณ์ใหม่"
สมรรถภาพในการคำนวณ
โดยประวัติแล้ว เชื่อว่า นิวรอนเป็นชิ้นส่วนที่ไม่ซับซ้อนและสมรรถภาพในการคำนวณของสมองมาจากการมีนิวรอนเป็นจำนวนมาก และจริง ๆ แล้ว งานศึกษาในเรื่องปัญญาประดิษฐ์ได้ดำเนินไปตามแนวทางนี้ แต่ปัจจุบันนี้ชัดเจนแล้วว่า นิวรอนเดี่ยว ๆ สามารถทำการคำนวณที่ซับซ้อน
แหล่งข้อมูลอื่น
- Kandel, Eric R.; Schwartz, James H.; Jessell, Thomas M.; Siegelbaum, Steven A.; Hudspeth, A.J. (2013). Principles of Neural Science Fifth Edition. United State of America: McGraw-Hill. ISBN 978-0-07-139011-8.
- Bullock, T.H., Bennett, M.V.L., Johnston, D., Josephson, R., Marder, E., Fields R.D. 2005. The Neuron Doctrine, Redux, Science, V.310, p. 791-793.
- Ramón y Cajal, S. 1933 Histology, 10th ed., Wood, Baltimore.
- Richard S. Snell: Clinical neuroanatomy (Lippincott Williams & Wilkins, Ed.6th 2006) Philadelphia, Baltimore, New York, London. ISBN 978-963-226-293-2
- Roberts A., Bush B.M.H. 1981. Neurones Without Impulses. Cambridge University Press, Cambridge.
- Peters, A., Palay, S.L., Webster, H, D., 1991 The Fine Structure of the Nervous System, 3rd ed., Oxford, New York
เว็บไซต์
- Neurobiology ที่เว็บไซต์ Curlie
- http://www.uppicth.com/bio.html
- IBRO (International Brain Research Organization) เก็บถาวร 2013-04-25 ที่ เวย์แบ็กแมชชีน. Fostering neuroscience research especially in less well-funded countries.
- NeuronBank an online neuromics tool for cataloging neuronal types and synaptic connectivity.
- High Resolution Neuroanatomical Images of Primate and Non-Primate Brains.
- The Department of Neuroscience at Wikiversity, which presently offers two courses: Fundamentals of Neuroscience and Comparative Neuroscience.
- NIF Search - Neuron เก็บถาวร 2015-01-22 ที่ เวย์แบ็กแมชชีน via the Neuroscience Information Framework
- Cell Centered Database - Neuron เก็บถาวร 2011-08-13 ที่ เวย์แบ็กแมชชีน
- Complete list of neuron types according to the Petilla convention, at NeuroLex.
- NeuroMorpho.Org an online database of digital reconstructions of neuronal morphology.
- Immunohistochemistry Image Gallery: Neuron เก็บถาวร 2011-10-08 ที่ เวย์แบ็กแมชชีน
- Khan Academy: Anatomy of a neuron
- Neuron images