
| เซลล์รับแสง (Photoreceptor Cell) | |
|---|---|
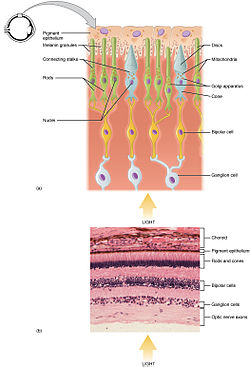 ส่วนประกอบต่าง ๆ ของเซลล์รูปแท่งและเซลล์รูปกรวย ซึ่งเป็นเซลล์ 2 ประเภทใน 3 ประเภทที่ไวแสงในจอประสาทตา
| |
| ตัวระบุ | |
| MeSH | D010786 |
| นิวโรเล็กซ์ ID | sao226523927 |
| FMA | 85613 86740, 85613 |
| ศัพท์ทางกายวิภาคของประสาทกายวิภาคศาสตร์ | |
เซลล์รับแสง (อังกฤษ: photoreceptor cell) เป็นเซลล์ประสาท (นิวรอน) พิเศษในจอประสาทตาที่มีสมรรถภาพในการถ่ายโอนแสงไปเป็นพลังประสาท ความสำคัญทางชีวภาพของเซลล์รับแสงก็คือความสามารถในการแปลงแสงที่เห็นได้ไปเป็นสัญญาณที่สามารถเร้ากระบวนการต่าง ๆ ทางชีวภาพ จะกล่าวให้ชัดเจนกว่านี้ก็คือ มีโปรตีนหน่วยรับแสงในเซลล์ที่ดูดซึมโฟตอน ซึ่งนำไปสู่ความเปลี่ยนแปลงในความต่างศักย์ของเยื่อหุ้มเซลล์
เซลล์รับแสงแบบคลาสิกก็คือเซลล์รูปแท่งและเซลล์รูปกรวย แต่ละอย่างล้วนแต่ให้ข้อมูลที่ใช้ในระบบการมองเห็นเพื่อสร้างแบบจำลองของโลกภายนอกที่เห็นทางตา เซลล์รูปแท่งนั้นบางกว่าเซลล์รูปกรวย และมีความกระจัดจายไปในจอประสาทตาที่แตกต่างกัน แม้ว่า กระบวนการเคมีที่ถ่ายโอนแสงไปเป็นพลังประสาทนั้นคล้ายคลึงกัน มีการค้นพบเซลล์รับแสงประเภทที่สามในช่วงคริสต์ทศวรรษ 1990 ซึ่งก็คือ photosensitive retinal ganglion cell เป็นเซลล์ที่ไม่ได้มีส่วนให้เกิดการเห็นโดยตรง แต่เชื่อกันว่า มีส่วนช่วยในระบบควบคุมจังหวะรอบวัน (circadian rhythms) และปฏิกิริยาปรับรูม่านตาแบบรีเฟล็กซ์
เซลล์รูปแท่งและเซลล์รูปกรวยมีหน้าที่แตกต่างกัน คือ เซลล์รูปแท่งไวแสงเป็นพิเศษ มีปฏิกิริยาต่อโฟตอนเพียงแค่ 6 อนุภาค ดังนั้น ในที่มีระดับแสงต่ำ การเห็นเกิดจากสัญญาณที่มาจากเซลล์รูปแท่งเท่านั้น ซึ่งอธิบายว่า ทำไมเราจึงไม่สามารถเห็นภาพสีได้ในที่สลัว ซึ่งก็คือเพราะมีแต่เซลล์รูปแท่งเท่านั้นที่ทำงานได้ในระดับแสงนั้น และเซลล์รูปกรวยเป็นส่วนที่ทำให้เกิดการเห็นภาพสี
ส่วนเซลล์รูปกรวยต้องใช้แสงระดับที่สูงกว่ามาก (คือต้องมีโฟตอนมากระทบมากกว่า) ก่อนที่จะเกิดการทำงาน ในมนุษย์ มีเซลล์รูปกรวยสามประเภท จำแนกโดยการตอบสนองต่อความยาวคลื่นแสงที่ต่าง ๆ กัน การเห็นสี (ในภาพ) เป็นการประมวลผลจากสัญญาณที่มาจากเซลล์รูปกรวยสามประเภทเหล่านี้ โดยน่าจะผ่านกระบวนการ opponent process เซลล์รูปกรวยสามอย่างนี้ตอบสนอง (โดยคร่าว ๆ) ต่อแสงที่มีความยาวคลื่นขนาดสั้น (S) ขนาดกลาง (M) และขนาดยาว (L) ให้สังเกตว่า การยิงสัญญาณของเซลล์รับแสงนั้นขึ้นอยู่เพียงกับจำนวนโฟตอนที่ได้รับเท่านั้น (กำหนดโดยทฤษฎี principle of univariance) ส่วนการตอบสนองที่ต่าง ๆ กันของเซลล์รูปกรวยขึ้นอยู่กับความเป็นไปได้ของโปรตีนรับแสงของเซลล์ที่จะดูดซึมแสงที่ความยาวคลื่นนั้น ๆ ยกตัวอย่างเช่น เซลล์รูปกรวยแบบ L มีโปรตีนรับแสงที่ดูดซึมแสงที่มีความยาวคลื่นขนาดยาว (หรือออกสีแดง ๆ) แม้ว่า แสงที่มีความยาวคลื่นสั้นกว่าอาจจะทำให้เกิดการตอบสนองในระดับเดียวกัน แต่จะต้องเป็นแสงที่สว่างกว่ามาก
จอประสาทตามมนุษย์มีเซลล์รูปแท่งประมาณ 120 ล้านเซลล์ และมีเซลล์รูปกรวยประมาณ 6 ล้านเซลล์ สัตว์ต่าง ๆ สปีชีส์มีอัตราส่วนของเซลล์รูปแท่งและเซลล์รูปกรวยที่แตกต่างกัน ขึ้นอยู่กับว่า เป็นสัตว์กลางวันหรือสัตว์กลางคืน นอกจากเซลล์รูปแท่งและเซลล์รูปกรวยแล้ว ยังมี retinal ganglion cell (ตัวย่อ RGC) ประมาณ 1.5 เซลล์ในมนุษย์ และมี 1-2% ที่ไวแสง
บทความนี้กล่าวถึงเซลล์รับแสงของสัตว์มีกระดูกสันหลัง เซลล์รับแสงของสัตว์ไม่มีกระดูกสันหลัง เช่นแมลงและมอลลัสกามีความแตกต่างจากสัตว์มีกระดูกสันหลังทั้งในโครงสร้างและในกระบวนการเคมีชีวภาพ
กายวิภาค
![กายวิภาคขอของเซลล์รูปแท่ง [10]](http://upload.wikimedia.org/wikipedia/commons/thumb/b/bb/Rod%26Cone.jpg/200px-Rod%26Cone.jpg)
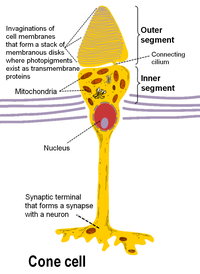
ในมนุษย์ เซลล์รูปแท่งและเซลล์รูปกรวยอยู่ที่ส่วนนอกสุดของจอประสาทตา (ด้านหลังสุดของตา) และมีโครงสร้างโดยพื้นฐานที่คล้ายคลึงกัน ส่วนที่ไกลจากสมองที่สุด เป็นส่วนสุดของแอกซอน (axon terminal) ซึ่งปล่อยสารสื่อประสาท "กลูตาเมต" ไปสู่ bipolar cell ส่วนเถิบต่อไปทางสมองเป็นตัวเซลล์ ซึ่งมีออร์แกเนลล์ของเซลล์ ส่วนเถิบต่อไปเรียกว่า inner segment ซึ่งเป็นส่วนมีหน้าที่พิเศษเต็มไปด้วยไมโทคอนเดรีย หน้าที่หลักของส่วนนี้ก็คือผลิต ATP ใช้เป็นพลังงานสำหรับการปั้มโซเดียมโพแทสเซียมผ่านเยื่อหุ้มเซลล์ ส่วนสุดท้ายที่ใกล้สมองที่สุดเรียกว่า outer segment ซึ่งเป็นส่วนที่ดูดซึมแสง ส่วนนี้จริง ๆ แล้วเป็นซีเลียดัดแปลง ประกอบด้วยจาน (disc) เต็มไปด้วยหน่วยรับความรู้สึกประเภท opsin ซึ่งเป็นโปรตีนที่ดูดซึมโฟตอน และเต็มไปด้วยประตูโซเดียมเปิดปิดโดยความต่างศักย์ (voltage-gated sodium channel)
โปรตีนรับแสงที่อยู่ที่เยื่อหุ้มเซลล์ที่เรียกว่า opsin มีโมเลกุลรงควัตถุที่เรียกว่า retinal และในเซลล์รูปแท่ง โปรตีนนี้รวมกับรงควัตถุเรียกว่า rhodopsin ส่วนในเซลล์รูปรวย มี opsin หลายประเภทที่รวมตัวกับ retinal เป็นรงควัตถุที่เรียกว่า photopsin ซึ่งแบ่งออกเป็นสามประเภท และมีปฏิกิริยาต่อแสงความถี่ต่าง ๆ กัน เป็นความต่างที่ยังระบบการมองเห็นให้สามารถแยกแยะสีได้ หน้าที่ของเซลล์รับแสงก็คือการเปลี่ยนพลังงานแสงจากโฟตอนให้เป็นพลังงานที่ใช้สื่อสารได้ในระบบประสาท เป็นการแปลงถ่ายพลังงานที่เรียกว่า การถ่ายโอนสัญญาณ (signal transduction)
ส่วนโปรตีน opsin ที่อยู่ใน retinal ganglion cell ไวแสง มีบทบาทในการตอบสนองแบบรีเฟล็กซ์ของสมองและร่างกายต่อแสง เช่นการควบคุมจังหวะรอบวัน (circadian rhythm) รีเฟล็กซ์รูม่านตา (pupillary reflex) และการตอบสนองประเภทอื่น ๆ ที่ไม่ได้ทำให้เกิดการเห็น เป็นโปรตีนที่เรียกว่า melanopsin ซึ่งมีน้อยในสัตว์มีกระดูกสันหลัง เป็นโปรตีนที่มีหน้าที่คล้ายกับ opsin ในสัตว์ไม่มีกระดูกสันหลัง เป็นประเภทหนึ่งของ opsin ซึ่งเป็นหน่วยรับความรู้สึกแบบ G-protein-coupled receptor ประเภท retinylidene protein
เมื่อแสงสว่างทำปฏิกิริยากับระบบส่งสัญญาณของ melanopsin, retinal ganglion cell (ตัวย่อ RGC) ที่ประกอบด้วย melanopsin ก็จะปล่อยพลังประสาทผ่านแอกซอนไปยังเป้าหมายเฉพาะต่าง ๆ ในสมอง รวมทั้งนิวเคลียส olivary pretectal nucleus (ศูนย์ที่มีหน้าที่ควบคุมรูม่านตา), lateral geniculate nucleus (ตัวย่อ LGN) และนิวเคลียส suprachiasmatic nucleus ของไฮโปทาลามัส (ซึ่งทำหน้าที่เป็นนาฬิกาหลักของระบบจังหวะรอบวัน) ตัว RGC ที่ประกอบด้วย melanopsin เชื่อกันว่ามีออกฤทธิ์ต่อนิวเคลียสเป้าหมาย โดยปล่อยสารสื่อประสาทกลูตาเมต และ pituitary adenylate cyclase activating polypeptide (PACAP) ที่ไซแนปส์
มนุษย์



จอประสาทตามนุษย์มีเซลล์รูปกรวยประมาณ 6 ล้านตัวและเซลล์รูปแท่ง 120 ล้านตัว ข้อมูลที่ส่งไปจากเซลล์ประสาททั้งสองไปรวมกันที่เซลล์ RGC และ bipolar cell เพื่อการแปลผลก่อนที่จะส่งต่อไปยัง lateral geniculate nucleus ในทาลามัส ส่วนตรงกลางขอจอประสาทตา (คือส่วนที่ตรงกับตรงกลางของเลนส์ตา) เป็นรอยบุ๋มจอตา (fovea) ซึ่งมีแต่เซลล์รูปกรวย และเป็นส่วนที่ทำให้เกิดการเห็นที่ชัดที่สุดที่ละเอียดที่สุด ส่วนที่เหลือของเรตินามีทั้งเซลล์รูปแท่งและเซลล์รูปกรวยอยู่ปนกันไป แต่ไม่มีเซลล์รับแสงอะไร ๆ ที่จุดบอด ซึ่งเป็นเขตที่ใยประสาทของ RGC รวมตัวเป็นเส้นประสาทตา (optic nerve) แล้วออกจากลูกตา
โปรตีนรับแสงของเซลล์รูปกรวยสามประเภทแตกต่างกันในความไวต่อโฟตอนของแสงมีความยาวคลื่นต่าง ๆ กัน (ดูรูป) เพราะว่า เซลล์รูปกรวยตอบสนองทั้งต่อความยาวคลื่น (wavelength) และทั้งต่อความเข้ม (intensity) ของแสง จึงต้องวัดความไวต่อความยาวคลื่นโดยอัตราการตอบสนองสัมพัทธ์ (relative rate) ของเซลล์ เมื่อค่าความเข้มแสงคงที่ และค่าความยาวคลื่นเป็นตัวแปร แล้วจึงจะได้ค่าอนุมานของ absorbance (การดูดกลืนแสง) กราฟที่เห็นแสดงค่าดูดกลืนแสงโดยแจกแจงทำให้เป็นบรรทัดฐานที่ 100 (normalized) ยกตัวอย่างเช่น เซลล์รูปกรวย S (ที่ดูดซึมแสงสั้น) ตอบสนองในระดับสูงสุดที่ความยาวคลื่น 420 นาโนเมตร ซึ่งบอกเราว่า เซลล์ชนิดนั้นมีความเป็นไปได้ที่จะดูดกลืนโฟตอนที่ 420 นาโนเมตรสูงกว่าความยาวคลื่นอื่น ๆ แต่ว่า ถ้าแสงมีความยาวคลื่นต่างจากระดับนั้น เช่นที่ 480 นาโนเมตร แต่ว่ามีระดับแสงที่เข้มขึ้น เซลล์นั้นก็จะตอบสนองในระดับเดียวกัน ดังนั้น เส้นโค้งในกราฟอาจจะทำให้เข้าใจผิดได้ เพราะว่า เซลล์รูปกรวยจริง ๆ ไม่สามารถตรวจจับสีโดยเพียงลำพัง แต่ว่า การเห็นสีจะต้องอาศัยการเปรียบเทียบสัญญาณจากเซลล์รูปกรวยประเภทต่าง ๆ
การถ่ายโอนแสง
กระบวนการถ่ายโอนแสง (phototransduction) นั้นเกิดขึ้นที่จอประสาทตา ซึ่งประกอบด้วยเซลล์ประสาทเป็นชั้น ๆ เซลล์รับแสง คือเซลล์รูปแท่งและเซลล์รูปกรวย อยู่ในชั้นในสุด ส่วนชั้นกลาง ๆ มี bipolar cell ซึ่งรับสัญญาณประสาทมาจากเซลล์รับแสงแล้วส่งข้อมูลต่อไปยังชั้นนอกสุดของจอประสาทตา ซึ่งเป็นชั้นที่ retinal ganglion cell (ตัวย่อ RGC) ประมวลผลสัญญาณแล้วจึงส่งต่อไปให้สมอง โดยส่งผ่านแอกซอนของ RGC ที่รวมตัวกันเป็นเส้นประสาทตาแล้วออกจากตาผ่านช่องในเรตินาซึ่งอยู่ที่จุดบอด
การทำงานของเซลล์รับแสงเป็นแบบ hyperpolarization (ไม่เหมือนเซลล์ประสาททั่ว ๆ ไปซึ่งทำงานแบบ depolarization) คือในขณะที่ไม่มีสิ่งเร้า เซลล์จะอยู่ในภาวะ depolarization และจะปล่อยสารสื่อประสาท "กลูตาเมต" ออกมาอย่างต่อเนื่อง ในที่มืด เซลล์มี cyclic guanosine 3'-5' monophosphate (cGMP) ในระดับความเข้มข้นสูง มีผลเป็นการเปิดประตูไอออน (ion channel) ให้โซเดียม (และแคลเซียมแม้จะไม่เป็นตัวหลัก) เข้ามาในเซลล์ได้ ชาร์จไอออนขั้วบวกที่ไหลเข้ามาในเซลล์เปลี่ยนความต่างศักย์ของเซลล์ก่อให้เกิด depolarization ซึ่งส่งผลให้เกิดการหลั่งสารสื่อประสาทกลูตาเมต ซึ่งอาจจะมีผลเป็น hyperpolarization หรือ depolarization สำหรับเซลล์ที่รับสัญญาณสืบ ๆ ไป
เมื่อแสงวิ่งเข้ามากระทบสารรงควัตถุไวแสงที่อยู่ในเซลล์รับแสง สารรงควัตถุนั้นจะเปลี่ยนรูปร่าง สารรงควัตถุนั้นเรียกว่า iodopsin หรือ rhodopsin และประกอบด้วยโปรตีนขนาดใหญ่เรียกว่า opsin (ซึ่งอยู่ที่เยื่อหุ้มเซลล์) ซึ่งเชื่อมอยู่กับกลุ่มโมเลกุล prosthetic group ที่ยึดอยู่ด้วยกันผ่านพันธะโคเวเลนซ์ รวมกันเป็นโมเลกุลอินทรีย์ที่เรียกว่า retinal (ซึ่งเป็นรูปแบบหนึ่งของวิตามินเอ) retinal มีสภาพเป็น 11-cis-retinal เมื่ออยู่ในที่มืด และเมื่อมีการกระตุ้นจากแสงก็จะเปลี่ยนสภาพไปเป็น all-trans-retinal ความเปลี่ยนแปลงทางโครงสร้างนี้จะเริ่มการทำงานของโปรตีนควบคุมเรียกว่า transducin ซึ่งเริ่มการทำงานของเอนไซม์ cGMP phosphodiesterase ซึ่งสลาย cGMP ออกเป็น 5'-GMP การริวดิวซ์ cGMP อย่างนี้ทำให้ประตูไอออนปิด ห้ามการไหลเข้าของไอออนบวก ซึ่งมีผลให้เซลล์เปลี่ยนสภาพเป็น hyperpolarization แล้วหยุดการปล่อยสารสื่อประสาท ขั้นตอนต่าง ๆ ทั้งหมดเริ่มต้นตั้งแต่แสงมากระทบที่เรตินา ซึ่งในที่สุดมีผลเป็นการตอบสนองของระบบรับรู้ เรียกว่า visual phototransduction
Dark current
เมื่อไม่มีสิ่งเร้า (ในที่มืด) ประตูไอออนที่เปิดปิดโดย cyclic-nucleotide ในส่วน outer segment ของเซลล์รับแสง จะเปิดอยู่เพราะว่า cyclic GMP (cGMP) ยึดอยู่กับประตูนั้น ดังนั้น ไอออนบวก (คือ ไอออนโซเดียม) ก็จะไหลเข้ามาในเซลล์รับแสง ทำให้เกิดภาวะ depolarization ในระดับ −40 mV (ในขณะที่ความต่างศักย์ในระดับพัก คือ resting potential ในเซลล์ประเภทอื่น ๆ อยู่ที่ −65 mV) กระแสไฟฟ้าที่มีผลเป็นสภาพ depolarization นี้เรียกว่า dark current (กระแสไฟฟ้าในที่มืด)
Signal transduction pathway

กระบวนการถ่ายโอนสัญญาณ (signal transduction pathway) เป็นกลไกที่พลังงานของโฟตอนเริ่มการทำงานของเซลล์มีผลเป็นการเปลี่ยนความต่างศักย์ (hyperpolarization) โดยที่สุด ซึ่งก็จะมีผลเป็นการส่งหรือการห้ามการส่งสัญญาณประสาท ซึ่งส่งต่อไปยังสมองผ่านเส้นประสาทตา กระบวนการถ่ายโอนสัญญาณของเซลล์รับแสงในสัตว์มีกระดูกสันหลังมีดังนี้
- rhodopsin หรือ iodopsin ที่เยื่อหุ้ม disc ใน outer segment ของเซลล์ดูดกลืนโฟตอน มีผลให้ Schiff base cofactor ของ retinal เปลี่ยนแบบจาก cis-form เป็น trans-form ทำให้ retinal เปลี่ยนรูปร่างไป
- ผลที่ตามมาก็คือความเปลี่ยนรูปไปเป็นสารต่าง ๆ ที่ไม่มีเสถียรภาพไปตามลำดับ แต่ตัวสุดท้ายจะสร้างพันธะที่มีกำลังกับ G protein ที่เยื่อหุ้มที่มีชื่อว่า transducin และจะเริ่มปฏิกิริยาของโปรตีนนี้ นี้เป็นการขยายสัญญาณในระดับแรก เพราะว่า rhodopsin แต่ละหน่วยที่เกิดการทำงานเพราะแสงจะนำไปสู่การทำงานของ transducin 100 หน่วย
- transducin แต่ละหน่วยจะเริ่มการทำงานของเอนไซม์ cGMP-specific phosphodiesterase (PDE)
- PDE จะเป็นตัวเร่งปฏิกิริยาการสลายตัวด้วยน้ำ (hydrolysis) ของ cGMP ไปเป็น 5' GMP ซึ่งเป็นการขยายสัญญาณในระดับที่สอง เพราะว่า PDE แต่ละหน่วยจะก่อให้เกิดการสลายตัวด้วยน้ำใน cGMP 1,000 โมเลกุล
- ระดับความเข้มข้นสุทธิของ cGMP ที่อยู่ในเซลล์จะลดลง (เพราะมีการแปลง cGMP ไปเป็น 5' GMP โดย PDE) มีผลเป็นการปิดประตูไอออน Na+ ที่เปิดปิดด้วย cyclic nucleotide (cyclic nucleotide-gated Na+ ion channel) ซึ่งอยู่ที่เยื่อหุ้มของ outer segment ของเซลล์รับแสง
- และดังนั้น ไอออนโซเดียมจะไม่สามารถไหลเข้ามาในเซลล์อีกต่อไป และเยื่อหุ้ม outer segment ของเซลล์รับแสงก็จะเปลี่ยนภาวะไปเป็น hyperpolarization เพราะว่าประจุไฟฟ้าภายในเยื่อหุ้มมีขั้วลบเพิ่มขึ้น
- ความเปลี่ยนแปลงความต่างศักย์ของเยื่อหุ้มเซลล์ก็จะมีผลเป็นการปิดประตูแคลเซียมซึ่งเปิดปิดโดยความต่างศักย์ และนำไปสู่การระงับการไหลเข้าของไอออนแคลเซียมเข้ามาในเซลล์ และดังนั้น ระดับความเข้มของไอออนแคลเซียมภายในเซลล์ก็จะลดลง
- เมื่อระดับความเข้มของไอออนแคลเซียมภายในเซลล์ลดลง เซลล์ก็จะปล่อยสารสื่อประสาทกลูตาเมตผ่านกระบวนการ exocytosis ไปยัง bipolar cell ลดลง (ซึ่งอาจจะมีผลเป็นการกระตุ้นหรือการยับยั้ง bipolar cell ที่อยู่หลังไซแนปส์)
- การลดระดับลงของการปล่อยกลูตาเมตจะทำให้ bipolar cell บางกลุ่มเปลี่ยนสภาพเป็น depolarization และบางกลุ่มจะเปลี่ยนสภาพเป็น hyperpolarization ขึ้นอยู่กับหน่วยรับความรู้สึกที่มีปฏิกิริยาต่อสารสื่อประสาทหลังไซแนปส์
ดังนั้น ให้สังเกตว่า ทั้งเซลล์รูปแท่งเซลล์รูปกรวยลดระดับการปล่อยสารสื่อประสาทเมื่อมีการกระตุ้นโดยแสง ซึ่งอาจจะกระตุ้น (มีผลเป็น depolarizatioin) หรือยับยั้ง (มีผลเป็น hyperpolarization) ใน bipolar cell ที่เซลล์รับแสงเชื่อมไซแนปส์ ขึ้นอยู่กับหน่วยรับความรู้สึกของ bipolar cell ความยืดหยุ่นได้อย่างนี้เป็นส่วนที่ขาดไม่ได้ในการสร้างโครงสร้างแบบ on-centre และ off-centre
เซลล์ส่วน inner segment ให้สาร ATP เพื่อเป็นพลังงานกับการปั้มโซเดียมโพแทสเซียม ซึ่งเป็นกระบวนการที่สำคัญเพื่อตั้งสภาพเริ่มต้นของ outer segment ขึ้นใหม่โดยสูบไอออนโซเดียมที่ไหลเข้ามาในเซลล์ออกไปนอกเซลล์
แม้ว่าเซลล์รับแสงจะเป็นเซลล์ประสาท แต่เป็นเซลล์ประสาทที่ไม่ส่งศักยะงาน (action potential) ยกเว้นแต่ RGC ไวแสง (photosensitive ganglion cell) ซึงมีหน้าที่หลักในการควบคุมจังหวะรอบวัน (circadian rhythms), melatonin, และการปรับรูม่านตา
ข้อได้เปรียบ
การถ่ายโอนแสงในเซลล์รับแสงไม่เหมือนใคร คือ ตัวกระตุ้น (ในที่นี่คือแสง) กลับลดระดับการตอบสนองของเซลล์ (คืออัตราการยิงสัญญาณ) ซึ่งเป็นการตอบสนองที่ไม่ทั่วไปในระบบรับความรู้สึก ซึ่งโดยทั่ว ๆ ไปแล้ว ตัวกระตุ้นจะเพิ่มการตอบสนองของเซลล์ อย่างไรก็ดี กระบวนการเช่นนี้มีข้อได้เปรียบที่สำคัญหลายอย่าง
ข้อที่หนึ่ง เซลล์รับแสงมีสภาพ depolarization ในที่มืด ซึ่งก็หมายความว่า ไอออนโซเดียมมากมายจะไหลเข้ามาในเซลล์ ดังนั้น การเปิดปิดประตูโซเดียมที่เกิดขึ้นเองจะไม่มีผลต่อความต่างศักย์ของเยื่อหุ้มเซลล์ และการปิดประตูเป็นจำนวนมากที่เนื่องมาจากการดูดกลืนโฟตอนของแสงเท่านั้น ที่จะมีผลเป็นการส่งสัญญาณว่า มีแสงภายในลานสายตา ดังนั้น กระบวนการเช่นนี้จะปราศจากสัญญาณรบกวน
ข้อที่สอง มีการขยายสัญญาณในระดับสูงแบ่งเป็นสองขั้นในการถ่ายโอนแสง คือ สารรงควัตถุ (pigment) จะปลุกฤทธิ์ของ transducin เป็นจำนวนหลายโมเลกุล และ PDE แต่ละตัวจะก่อให้เกิดการสลายตัวของ cGMP หลายตัว เพราะมีการขยายสัญญาณอย่างนี้ การดูดกลืนโฟตอนเพียงแค่อนุภาคเดียวก็จะมีผลต่อความต่างศักย์เยื่อหุ้มเซลล์ ทำให้เกิดการส่งข้อมูลไปยังสมองว่า มีแสงอยู่ภายในลานสายตา นี่เป็นจุดแตกต่างที่สำคัญระหว่างเซลล์รูปแท่งและเซลล์รูปกรวยซึ่งล้วนแต่เป็นเซลล์รับแสง คือ เซลล์รูปแท่งไวแสงมากและสามารถรับรู้ถึงโฟตอนแม้เพียงอนุภาคเดียว ซึ่งไม่เหมือนกับเซลล์รูปกรวย ส่วนเซลล์รูปกรวยมีอัตราการขยายสัญญาณในระหว่างการถ่ายโอนสัญญาณที่รวดเร็วมาก ซึ่งไม่เหมือนเซลล์รูปแท่ง
ความต่างกันระหว่างเซลล์รูปกรวยและเซลล์รูปแท่ง
การเปรียบเทียบเซลล์รูปกรวยและเซลล์รูปแท่งต่อไปนี้ มาจากหนังสือ Principles of Neural Science (หลักประสาทวิทยาศาสตร์) ของ ศ. อีริก แคนเดิล ผู้ได้รับรางวัลโนเบลสาขาสรีรวิทยาหรือการแพทย์ในงานวิจัยเกี่ยวกับพื้นฐานทางสรีรภาพในการบันทึกความจำในนิวรอน
| เซลล์รูปแท่ง | เซลล์รูปกรวย |
|---|---|
| ให้เกิดการเห็นใต้แสงสลัว (scotopic vision) | ให้เกิดการเห็นใต้ใต้แสงเข้ม (photopic vision) |
| ไวแสงมาก แม้แต่ต่อแสงที่กระจัดกระจาย | ไม่ไวแสงมาก ไวต่อแสงตรง ๆ เท่านั้น |
| ถ้าเสียจะทำให้ตาบอดมืด | ถ้าเสียจะทำให้ตาบอดตามกฎหมาย |
| มองเห็นได้ไม่ชัด | มองเห็นได้ชัด มีรายละเอียดสูง |
| ไม่มีในรอยบุ๋มจอตา | หนาแน่นที่สุดในรอยบุ๋มจอตา |
| ตอบสนองต่อแสงช้าโดยเพิ่มขึ้นเรื่อย ๆ | ตอบสนองต่อแสงได้เร็ว สามารถเห็นภาพที่กำลังเปลี่ยนแปลงอย่างรวดเร็ว |
| มีสารรงควัตถุมากกว่า จึงสามารถเห็นในที่สลัว | มีสารรงควัตถุน้อยกว่า จึงต้องใช้แสงมากกว่าเพื่อจะเห็น |
| จาน (disk) ตั้งอยู่ภายในเยื่อหุ้ม ไม่ได้ยึดอยู่กับเยื่อหุ้มเซลล์โดยตรง | จานยึดไว้กับเยื่อหุ้มเซลล์ |
| มีประมาณ 120 ล้านเซลล์ในจอประสาทตาแต่ละข้าง | มีประมาณ 6 ล้านเซลล์ในจอประสาทตา |
| มีสารรงควัตถุประเภทเดียว | มีสารรงควัตถุ 3 ประเภทในมนุษย์ |
| ทำให้เกิดการเห็นปราศจากสี | ทำให้เกิดการเห็นภาพสี |
หน้าที่
เซลล์รับแสงไม่ได้ส่งข้อมูลเกี่ยวกับสีโดยตรง แต่ส่งสัญญาณว่ามีแสงอยู่ในลานสายตา
เซลล์รับแสงหนึ่ง ๆ จะตอบสนองต่อทั้งความยาวคลื่นและความเข้มของแสง ยกตัวอย่างเช่น การตอบสนองของเซลล์รับแสงหนึ่งต่อแสงสีแดงที่มีความเข้มระดับหนึ่ง อาจจะเหมือนกันกับการตอบสนองต่อแสงสีเขียวที่มีความเข้มอีกระดับหนึ่ง ดังนั้น การตอบสนองของเซลล์รับแสงหนึ่ง ๆ ไม่ได้บอกอย่างชัดเจนว่าเห็นสีอะไร
ดังนั้น เพื่อจะระบุสี ระบบการมองเห็นต้องเปรียบเทียบการตอบสนองของกลุ่มเซลล์รับแสงต่าง ๆ คือโดยเฉพาะแล้ว จากกลุ่มเซลล์รูปกรวย 3 ประเภทที่ดูดกลืนแสงที่มีความยาวคลื่นต่าง ๆ กัน และเพื่อที่จะระบุความเข้มแสง ระบบการมองเห็นต้องคำนวณว่ามีเซลล์รับแสงกี่ตัวที่กำลังทำการตอบสนอง นี้เป็นกลไกที่ทำให้เกิดการเห็นโดย 3 สีหลัก (trichromacy) ในทั้งมนุษย์และในสัตว์อื่น ๆ บางชนิด
พัฒนาการ
กระบวนการพัฒนาการเป็นเซลล์รูปแท่ง เซลล์รูปกรวยแบบ S และเซลล์รูปกรวยแบบ M เริ่มจากแฟกเตอร์การถอดรหัส (transcription factor) หลายอย่าง รวมทั้ง RORbeta, OTX2, NRL, CRX, NR2E3 และ TRbeta2. โดยทั่วไปแล้ว เซลล์เบื้องต้นจะเจริญเป็นเซลล์รูปกรวยแบบ S โดยปริยาย แต่เหตุการณ์การถอดรหัสต่าง ๆ สามารถทำให้เจริญเป็นเซลล์รูปแท่งหรือเซลล์รูปกรวยแบบ M แม้ว่า เซลล์รูปกรวยแบบ L จะมีอยู่ในสัตว์อันดับวานร แต่ว่า การพัฒนาการของเซลล์ไม่เป็นที่รู้จักกันดีเนื่องจากว่าสัตว์ทดลองที่ใช้เป็นสัตว์อันดับสัตว์ฟันแทะที่ไม่มีเซลล์ชนิดนี้ มี 5 ขั้นตอนในการพัฒนาเป็นเซลล์รับแสง คือ
- การแพร่ขยายของ retinal progenitor cell (เซลล์บรรพบุรุษ, เซลล์เบื้องต้น, ตัวย่อ RPC) ที่สามารถพัฒนาเป็นเซลล์ได้หลายชนิด
- restriction of competence of RPCs;
- การกำหนดชะตาของเซลล์ (cell fate specification)
- การแสดงออกของยีนเป็นเซลล์รับแสง
- และขั้นสุดท้ายคือการงอกออกของแอกซอน การตั้งไซแนปส์ และการเจริญของส่วน outer segment
ในส่วนเบื้องต้น Notch signaling pathway จะทำให้เกิดการหมุนเวียนของเซลล์บรรพบุรุษ (progenitor cycling) เพราะฉะนั้น เซลล์รับแสงตั้งต้น (Photoreceptor precursor) จะเกิดขึ้นได้ก็เมื่อเกิดการยับยั้ง Notch signaling pathway และมีการทำงานในระดับที่สูงขึ้นของกระบวนการต่าง ๆ รวมทั้งกลุ่มยีน achaete-scute complex (หรือ achaete-scute homologue) ส่วน แฟกเตอร์ OTX2 กำหนดชะตาเซลล์ให้เป็นเซลล์รับแสง ส่วน CRX กำหนดยีนของเซลล์รับแสงที่จะเกิดการแสดงออก ส่วนการแสดงออกของ NRL กำหนดชะตาของเซลล์ให้เป็นเซลล์รูปแท่ง ส่วน NR2E3 ทำการกำหนดชะตาเป็นเซลล์รูปแท่งให้มั่นคงยิ่งขึ้นโดยห้ามการแสดงออกของยีนเซลล์รูปกรวย ส่วนแฟกเตอร์ RORbeta ใช้ในการพัฒนาทั้งของเซลล์รูปแท่งและทั้งของเซลล์รูปกรวย ส่วน TRbeta2 กำหนดชะตาให้เป็นเซลล์รูปกรวยแบบ M ถ้าเอาออกแฟกเตอร์ใดแฟกเตอร์หนึ่งที่กล่าวมาแล้ว ชะตาโดยปริยายของเซลล์ก็คือเซลล์รูปกรวยแบบ S ให้สังเกตว่าขั้นตอนต่าง ๆ เหล่านี้เกิดขึ้นในช่วงเวลาต่าง ๆ กันสำหรับสัตว์สปีชีส์ต่าง ๆ กัน ผ่านกระบวนการซับซ้อนต่าง ๆ ที่มีผลเป็นลักษณะทางพันธุกรรมที่แตกต่างกัน ถ้าตัวควบคุมกระบวนการเหล่านี้เกิดความเสียหาย โรค retinitis pigmentosa จอตาเสื่อมที่จุดภาพชัด (macular degeneration) และความผิดปกติทางตาอย่างอื่นอาจเกิดขึ้น
การส่งสัญญาณ
เซลล์รับแสงส่งข้อมูลเกี่ยวกับการดูดกลืนโฟตอนโดยลดระดับการปล่อยสารสื่อประสาทกลูตาเมตที่ไซแนปส์ที่เชื่อมกับ bipolar cell เพราะว่ามีสภาพเป็น depolarization ในที่มืด คือจะมีการปล่อยกลูตาเมตเป็นจำนวนมากไปยัง bipolar cell ในที่มืด การดูดกลืนโฟตอนหนึ่ง ๆ จะมีผลให้เกิดสภาพ hyperpolarization ในเซลล์รับแสง และดังนั้นจึงมีผลเป็นการลดระดับการปล่อยกลูตาเมตเข้าไปยังไซแนปส์ที่เชื่อมต่อกับ bipolar cell
แม้ว่า เซลล์รูปแท่งและเซลล์รูปกรวยทุก ๆ ตัวจะปล่อยสารสื่อประสาทประเภทเดียวกันคือกลูตาเมต แต่ว่าผลที่เกิดขึ้นจากสารสื่อประสาทเดียวกันมีผลต่าง ๆ กันต่อ bipolar cell ต่าง ๆ กัน ขึ้นอยู่กับหน่วยรับความรู้สึก (receptor) ที่อยู่ที่เยื่อหุ้มเซลล์ของ bipolar cell ถ้ากลูตาเมตเกิดพันธะขึ้นกับ ionotropic receptor เซลล์นี้ก็จะมีสภาพเป็น depolarization ในที่มืดและดังนั้น จะมีสภาพเป็น hyperpolarization เมื่อมีแสงเพราะมีกลูตาเมตในระดับที่ลดลง ในนัยตรงกันข้าม ถ้ากลูตาเมตเกิดพันธะขึ้นกับ metabotropic receptor เซลล์นี้ก็จะมีสภาพเป็น hyperpolarization ในที่มืด และมีสภาพเป็น depolarization เมื่อมีแสงเพราะว่าระดับของกลูตาเมตลดลง
โดยทั่ว ๆ ไปแล้ว คุณสมบัติเช่นนี้จึงยังให้เซลล์กลุ่มหนึ่งเกิดการเร้าและอีกกลุ่มหนึ่งเกิดการยับยั้งเพราะแสง แม้ว่า เซลล์รับแสงทั้งหมดจะตอบสนองในลักษณะเดียวกันต่อแสง ความเป็นไปอันซับซ้อนเช่นนี้สำคัญและขาดไม่ได้ในการตรวจจับสี (color) ความเปรียบต่าง (contrast) และขอบ (edge)
ความซับซ้อนอื่น ๆ เกิดจากการเชื่อมต่อต่าง ๆ กันระหว่าง bipolar cell, horizontal cell, และ amacrine cell ในจอประสาทตา เป้าหมายที่สุดของการส่งสัญญาณซับซ้อนเช่นนี้ก็คือ กลุ่ม RGC ต่าง ๆ ซึ่งส่วนหนึ่งเป็นเซลล์ไวแสงเอง แต่ว่าจะใช้สารรงควัตถุไวแสง (photopigment) ประเภท melanopsin
RGC ไวแสง
ในปี ค.ศ. 1991 ฟอสเตอร์และคณะได้ค้นพบเซลล์รับแสงที่ไม่ใช่เซลล์รูปแท่งและเซลล์รูปกรวยในตาหนู ที่สื่อการควบคุมจังหวะรอบวัน (circadian rhythm) เซลล์รับแสงที่เรียกว่า intrinsically photosensitive retinal ganglion cells (ตัวย่อ ipRGC แปลว่า RGC ไวแสงโดยธรรมชาติ) เหล่านี้ เป็นส่วนย่อย ๆ (~1–3%) ของ RGC ที่อยู่ในเรตินาชั้นด้านใน ซึ่งก็คืออยู่ด้านหน้า (ดูโครงสร้างในจอประสาทตา) ของเซลล์รูปแท่งและเซลล์รูปกรวยซึ่งอยู่ด้านนอกของเรตินา เซลล์ประสาทไวแสงเหล่านี้มีสารรงควัตถุไวแสงที่เรียกว่า melanopsin ที่ดูดกลืนแสงที่มีความยาวคลื่นในระดับสูงสุดที่ ~480 นาโนเมตร ซึ่งต่างจากเซลล์รูปแท่งและเซลล์รูปกรวย นอกจากหน้าที่เกี่ยวกับจังหวะรอบวันและพฤติกรรมอย่างอื่น ๆ แล้ว ipRGCs ยังมีบทบาทในการเริ่มรีเฟล็กซ์ปรับรูม่านตา (pupillary light reflex) อีกด้วย
ในปี ค.ศ. 2005 เด็นนิส เดซีย์และคณะได้แสดงในวงศ์ลิงโลกเก่าว่า RGC แบบยักษ์ (giant ganglion cell) ที่มีการแสดงออกของ melanopsin นั้น ส่งแอกซอนไปยัง lateral geniculate nucleus ในทาลามัส คือ ก่อนหน้านี้ ได้มีการค้นพบแต่แอกซอนที่ส่งไปยัง pre-tectal nucleus ในสมองส่วนกลาง และไปยัง suprachiasmatic nucleus ในไฮโปทาลามัสเท่านั้น ถึงกระนั้น หน้าที่เกี่ยวกับการเห็นของเซลล์นี้ยังไม่ชัดเจน (ถ้ามี)
ในปี ค.ศ. 2007 ฟาร์ฮาน ไซดี และคณะได้ตีพิมพ์งานวิจัยต้นฉบับที่ทำการทดลองในมนุษย์ที่ไม่มีเซลล์รูปแท่งไม่มีเซลล์รูปกรวย ต่อมา วารสาร Current Biology ได้ประกาศในปี ค.ศ. 2008 ในบทบรรณาธิการว่า มีการค้นพบเซลล์รับแสงในมนุษย์ที่ไม่ใช่เซลล์รูปแท่งไม่ใช่เซลล์รูปกรวยโดยมีหลักฐานเป็นข้อยุติจากการทดลองสำคัญในมนุษย์โดยไซดีและคณะ คือโดยเหมือนกับที่พบในสัตว์เลี้ยงลูกด้วยนมประเภทอื่น ๆ เซลล์ที่ไม่ใช่เซลล์รูปแท่งไม่ใช่เซลล์รูปกรวยที่ไวแสงในมนุษย์ก็คือ RGC ในเรตินาด้านใน ในงานวิจัยนี้ นักวิจัยได้ตามหาคนไข้ที่หายากที่โรคได้ทำลายเซลล์รูปกรวยและเซลล์รูปแท่งแล้วอย่างสิ้นเชิง แต่ไม่ได้ทำอะไรต่อ RGC แม้ว่าจะไม่มีเซลล์รูปแท่งหรือเซลล์รูปกรวย คนไข้ก็ยังมียังมีพฤติกรรมเกี่ยวกับจังหวะรอบวัน, melanopsin suppression, และรีเฟล็กซ์ปรับรูม่านตาที่เหลืออยู่ โดยมีระดับความไวแสงสอดคล้องกับการมีสารรงควัตถุไวแสง melanopsin และสมองก็ยังมีการเห็นแสงที่มีความถี่เท่านี้อีกด้วย
ในมนุษย์ RGC ไวแสงมีส่วนในการเห็น (conscious sight) และในหน้าที่อื่น ๆ ที่ไม่เกี่ยวกับการเห็นรวมทั้งที่เกี่ยวกับจังหวะรอบวัน พฤติกรรม และการปรับรูม่านตา เพราะว่า เซลล์เหล่านี้ตอบสนองโดยมากกับแสงสีน้ำเงิน จึงมีข้อเสนอว่าเซลล์เหล่านี้มีบทบาทใน mesopic vision (การเห็นในที่สลัวไม่ถึงกับมืด) งานของไซดีและคณะในมนุษย์ที่ไม่มีเซลล์รูปแท่งไม่มีเซลล์รูปกรวยยังเปิดทางให้กับงานทดลองเกี่ยวกับบทบาทของ RGC ไวแสงต่อการเห็นอีกด้วย คือ เกิดการพบว่า มีวิถีประสาทขนานกันที่มีหน้าที่เกี่ยวกับการเห็น วิถีหนึ่งเป็นวิถีคลาสสิกไปจากเซลล์รูปแท่งและเซลล์รูปกรวยที่อยู่ทางด้านนอกของเรตินา และอีกวิถีหนึ่งเป็นการตรวจจับระดับแสงสว่างแบบคร่าว ๆ ที่เกิดจากเรตินาด้านใน ซึ่งดูเหมือนจะเกิดการทำงานก่อนด้านนอก นอกจากนั้นแล้ว ระบบคลาสสิกยังป้อนข้อมูลให้กับระบบที่ค้นพบใหม่อีกด้วย และนักวิจัยฟอสตอร์ได้เสนอว่า ความสม่ำเสมอของสี (colour constancy) อาจจะเป็นหน้าที่สำคัญของระบบใหม่
เซลล์รับแสงนี้อาจสำคัญต่อการเข้าใจโรคต่าง ๆ รวมทั้งโรคที่ทำให้เกิดตาบอดทั่วโลกรวมทั้งต้อหิน ซึ่งเป็นโรคที่มีผลต่อ RGC และการศึกษาเซลล์รับแสงนี้ อาจทำให้สามารถค้นหาแนวทางใหม่ ๆ ในการบำบัดรักษาโรคที่ให้เกิดตาบอด การค้นพบเซลล์รับแสงใหม่ ๆ ในมนุษย์ และการศึกษาว่า เซลล์เหล่านี้มีบทบาทในการเห็นอย่างไร (ไม่ใช่บทบาทในพฤติกรรมที่ไม่เกี่ยวกับการเห็นอย่างอื่น ๆ) นี่แหละ จะมีผลกว้างขวางที่สุดในสังคมมนุษย์ แม้ว่า ความบกพร่องเกี่ยวกับจังหวะรอบวันก็เป็นเรื่องที่น่าสนใจในการแพทย์รักษาอีกอย่างหนึ่ง
ในปี ค.ศ. 2003 สตีเว็น ล็อกลีย์และคณะแสดงว่า แสงมีความยาวคลื่นที่ 460 นาโนเมตรสามารถลดระดับ melatonin (ซึ่งเป็นฮอร์โมนในเลือดที่สื่อจังหวะรอบวัน) เป็นระยะเวลายาวถึงสองเท่านานกว่าที่ความยาวคลื่น 555 นาโนเมตร งานวิจัยโดยมากบอกเป็นนัยว่า ระดับความยาวคลื่นที่ RGC ไวแสงตอบสนองมากที่สุดอยู่ระหว่าง 460-482 นาโนเมตร แต่ว่า งานวิจัยหลังจากนั้นโดยฟาร์ฮาน ไซดีและคณะในมนุษย์ที่ไม่มีเซลล์รูปแท่งไม่มีเซลล์รูปกรวยพบว่า แสงความเข้มสูงที่นำไปสู่การรับรู้อยู่ที่ความยาวคลื่น 481 นาโนเมตร ซึ่งก็หมายความว่า RGC ไวแสงนี้ ยังให้การเห็นขั้นพื้นฐานเกิดขึ้นได้ดีที่สุดในแสงสีน้ำเงิน
ดูเพิ่ม
-
Campbell, Neil A., and Reece, Jane B. (2002). Biology. San Francisco: Benjamin Cummings. pp. 1064–1067. ISBN 0-8053-6624-5.
{{cite book}}: CS1 maint: multiple names: authors list (ลิงก์) - Freeman, Scott (2002). Biological Science (2nd Edition). Englewood Cliffs, N.J: Prentice Hall. pp. 835–837. ISBN 0-13-140941-7.
- NIF Search – Photoreceptor Cell เก็บถาวร 2016-03-04 ที่ เวย์แบ็กแมชชีน via the Neuroscience Information Framework