
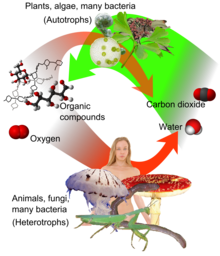
การหายใจระดับเซลล์ (อังกฤษ: cellular respiration) เป็นชุดปฏิกิริยาและกระบวนการทางเมแทบอลิซึมที่เกิดในเซลล์สิ่งมีชีวิตเพื่อเปลี่ยนแปลงพลังงานชีวเคมีจากสารอาหารเป็นอะดีโนซีนไตรฟอสเฟต (ATP) และปล่อยผลิตภัณฑ์ของเสียออกมา ปฏิกิริยาที่เกี่ยวข้องในการหายใจมีปฏิกิริยาแคแทบอลิซึมที่เกี่ยวข้องกับปฏิกิริยารีดอกซ์ (หมายถึง มีทั้งปฏิกิริยาออกซิเดชันและรีดักชัน) การหายใจเป็นหนึ่งในวิธีการหลักที่เซลล์จะได้รับพลังงานที่มีประโยชน์เพื่อเป็นเชื้อเพลิงการเปลี่ยนแปลงของเซลล์
สารอาหารซึ่งเซลล์สัตว์และพืชมักใช้ในการหายใจ มีน้ำตาล กรดอะมิโนและกรดไขมัน ตลอดจนตัวออกซิไดซ์ทั่วไป (ตัวรับอิเล็กตรอน) ในโมเลกุลออกซิเจน (O2) แบคทีเรียและอาร์เคียยังเป็นลิโธโทรฟ (lithotroph) คือ อาจหายใจได้โดยใช้โมเลกุลอนินทรีย์หลากชนิดเป็นตัวให้และรับอิเล็กตรอน เช่น กำมะถัน ไอออนโลหะ มีเทนและไฮโดรเจน สิ่งมีชีวิตซึ่งใช้ออกซิเจนเป็นตัวรับอิเล็กตรอนตัวสุดท้ายในการหายใจเรียกว่า สิ่งมีชีวิตต้องการออกซิเจน (aerobic) ส่วนสิ่งมีชีวิตที่ไม่ใช้ออกซิเจนเป็นตัวรับอิเล็กตรอนตัวสุดท้าย เรียกว่า สิ่งมีชีวิตไม่ต้องการออกซิเจน (anaerobic)
พลังงานซึ่งปลดปล่อยออกมาในการหายใจใช้ในการสังเคราะห์เอทีพีเพื่อเก็บพลังงานนี้ พลังงานที่เก็บในเอทีพีจากนั้นสามารถใช้เพื่อขับเคลื่อนขบวนการซึ่งอาศัยพลังงาน ได้แก่ ชีวสังเคราะห์ การเคลื่อนที่หรือการส่งโมเลกุลข้ามเยื่อหุ้มเซลล์
การหายใจแบบใช้ออกซิเจน
การหายใจแบบใช้ออกซิเจนต้องใช้ออกซิเจนเพื่อผลิตพลังงาน (ATP) แม้ว่าสารพวกคาร์โบไฮเดรต ลิพิดและโปรตีนจะสามารถผ่านขบวนการและใช้เป็นตัวทำปฏิกิริยาได้ทั้งหมด แต่ร่างกายมักใช้วิธีการสลายไพรูเวตในไกลโคไลสิส และกำหนดให้ไพรูเวตเข้าสู่ไมโทคอนเดรียเพื่อให้ถูกออกซิไดซ์ (oxideze) อย่างสมบูรณ์ในวัฏจักรเครปส์ ผลิตภัณฑ์ของขบวนการนี้คือ พลังงานในรูปของ ATP โดยฟอสโฟรีเลชันระดับซับสเตรต (substrate-level phosphorylation) NADH และ FADH2
| สมการปฏิกิริยาอย่างง่าย: | C6H12O6 (aq) + 6 O2 (g) → 6 CO2 (g) + 6 H2O (l) + ความร้อน |
| ΔG = -2880 kJ ต่อโมลของ C6H12O6 (กลูโคส) |
ค่า ΔG ที่ติดลบ แสดงว่า ปฏิกิริยานี้เกิดเองได้
ศักย์รีดิวซ์ของ NADH และ FADH2 ถูกแปลงไปเป็น ATP เพิ่มขึ้นอีกผ่านลูกโซ่ของการขนส่งอิเล็กตรอน (electron transport chain) โดยมีออกซิเจนเป็นตัวรับอิเล็กตรอนตัวสุดท้าย ATP ส่วนใหญ่ที่ผลิตจากการหายใจระดับเซลล์แบบใช้ออกซิเจนเป็นผลของออกซิเดทีฟฟอสโฟรีเลชัน ซึ่งทำงานได้โดยพลังงานที่ปลดปล่อยออกมาจากการสลายไพรูเวตนำไปใช้ในการปั๊มโปรตอนข้ามเยื่อหุ้ม เพื่อให้เกิดศักย์เคมิออสโมซิส (chemiosmosis) จากนั้น ศักย์เคมิออสโมซิสดังกล่าวจะถูกใช้ขับเคลื่อนเอทีพีซินเทส (ATP synthase) และสร้าง ATP จาก ADP และหมู่ฟอสเฟต ตำราชีววิทยามักระบุว่า กลูโคสหนึ่งโมเลกุลสามารถออกซิไดซ์ได้ ATP สูงสุด 38 โมเลกุลในการหายใจระดับเซลล์ (2 จากไกลโคไลสิส 2 จากวัฏจักรเครปส์ และ 34 จากลูกโซ่ของการขนส่งอิเล็กตรอน) ทว่า ในร่างกายแทบไม่เคยได้ผลลัพธ์สูงสุดดังกล่าวเลย เนื่องจากการสูญเสีย ซึ่งอาจเนื่องมาจากเยื่อหุ้มรั่ว เช่นเดียวกับค่าการขนส่งไพรูเวตและ ADP เข้าสู่เมตริกซ์ของไมโทคอนเดรียและปัจจุบันประเมินไว้ที่ราว 29 ถึง 30 ATP ต่อกลูโคส
เมแทบอลิซึมที่ใช้ออกซิเจนมีประสิทธิภาพสูงกว่าเมแทบอลิซึมแบบไม่ใช้ออกซิเจนถึง 15 เท่า (ซึ่งให้ ATP 2 โมเลกุลต่อ 1 โมเลกุลกลูโคส) ทั้งสองมีวิถีไกลโคไลสิสเริ่มต้นเหมือนกัน แต่เมแทบอลิซึมแบบใช้ออกซิเจนจะเกิดวัฏจักรเครปส์และออกซิเดทีฟฟอสโฟรีเลชันต่อ ซึ่งวัฏจักรเครปส์และออกซิเดทีฟฟอสโฟรีเลชันเกิดขึ้นในไมโทคอนเดรียในเซลล์ยูคาริโอต และไซโทพลาสซึมของเซลล์โปรคาริโอต
ไกลโคไลซิส
ไกลโคไลสิสเป็นวิถีเมแทบอลิซึมที่เกิดในไซโตซอลของเซลล์สิ่งมีชีวิตทุกชนิด วิถีดังกล่าวไม่ต้องอาศัยออกซิเจน จึงสามารถเกิดได้แม้ในสภาวะขาดออกซิเจน ขบวนการนี้เปลี่ยนกลูโคสหนึ่งโมเลกุลเป็นไพรูเวตสองโมเลกุล และสร้างพลังงานออกมาในรูปของ ATP สุทธิสองโมเลกุล แท้จริงแล้ว ในขบวนการดังกล่าวได้ ATP ออกมาสี่โมเลกุลต่อกลูโคส แต่มีการใช้ ATP สองโมเลกุลในระยะการเตรียม (preparatory phase) ฟอสโฟรีเลชันขั้นต้นของกลูโคสมีความจำเป็นในการทำให้โมเลกุลไม่เสถียรเพื่อแยกออกเป็นไพรูเวตสองโมเลกุล ระหว่างขั้นได้พลังงานคืน (pay-off phase) ของไกลโคไลสิส หมู่ฟอสเฟตสี่หมู่ถูกขนส่งไปยัง ADP โดยฟอสโฟรีเลชันระดับซับสเตรตเพื่อสร้าง ATP สี่โมเลกุล และมีการสร้าง NADH สองโมเลกุลเมื่อออกซิไดซ์ไพรูเวต สมการปฏิกิริยารวมเขียนได้ดังนี้
- กลูโคส + 2 NAD+ + 2 Pi + 2 ADP → 2 ไพรูเวต + 2 NADH + 2 ATP + 2 H+ + 2 H2O + ความร้อน
ออกซิเดทีฟดีคาร์บอกซิเลชันของไพรูเวต
ไพรูเวตถูกออกซิไดซ์เป็นอะซิติลโค เอ และคาร์บอนไดออกไซด์โดยไพรูวิกดีไฮโดรจิเนสคอมเพล็กซ์ (pyruvate dehydrogenase complex, PDC) ซึ่งประกอบด้วยเอนไซม์สามตัวและพบในไมโทคอนเดรียของเซลล์ยูคาริโอตและในไซโตซอลของโปรคาริโอต ในการเปลี่ยนไพรูเวตเป็นอะซิติลโค เอ จะได้ NADH และคาร์บอนไดออกไซด์อย่างละหนึ่งโมเลกุล ขั้นนี้ยังเป็นปฏิกิริยาเชื่อมโยงหรือปฏิกิริยาเปลี่ยนผ่าน ซึ่งเชื่อมระหว่างไกลโคไลสิสกับวัฏจักรเครปส์
วัฏจักรเครปส์
มีชื่อเรียกอื่นอีก ได้แก่ วัฏจักรกรดซิตริก หรือวัฏจักรกรดไตรคาร์บอกซิลิก เมื่ออยู่ในสภาวะที่ออกซิเจนเพียงพอ อะซิติลโค เอจะถูกสร้างขึ้นจากโมเลกุลไพรูเวตซึ่งได้มาจากไกลโคไลสิส เมื่อสร้างอะซิติลโค เอขึ้นมาแล้ว จะสามารถเกิดขบวนการต่อไปได้สองขบวนการ คือ การหายใจแบบใช้ออกซิเจนหรือไม่ใช้ออกซิเจน ในสภาวะออกซิเจนไม่เพียงพอ จะเกิดการหมักโมเลกุลไพรูเวต ส่วนในสภาวะที่ออกซิเจนเพียงพอ ไมโทคอนเดรียจะดำเนินการหายใจแบบใช้ออกซิเจน อะซิติลโค เอจะถูกนำเข้าสู่วัฏจักรเครปส์ในเมตริกซ์ของไมโทคอนเดรีย และถูกออกซิไดซ์จนได้เป็นคาร์บอนไดออกไซด์ ขณะที่ริวดิวซ์ NAD เป็น NADH ไปพร้อมกัน NADH สามารถถูกใช้สร้าง ATP เพิ่มอีกโดยลูกโซ่ของการขนส่งอิเล็กตรอน ในออกซิเดทีฟฟอสโฟรีเลชัน เพื่อที่จะออกซิไดซ์อย่างสมบูรณ์ให้เทียบเท่ากลูโคสหนึ่งโมเลกุล อะซิติลโค เอสองโมเลกุลต้องเกิดเมแทบอลิซึมโดยวัฏจักรเครปส์ วัฏจักรนี้ได้ผลิตภัณฑ์ของเสียออกมาสองอย่าง คือ น้ำและคาร์บอนไดออกไซด์
วัฏจักรเครปส์เป็นขบวนการ 8 ขั้นตอนที่มีเอนไซม์และโคเอนไซม์จำนวนมากเข้ามาเกี่ยวข้อง สารตัวกลางในวัฏจักร ได้แก่ ซิเตรด (citrate) ไอโซซิเตรด (isocitrate) แอลฟาคีโตกลูตาเรต (α-Ketoglutarate) ซักซินิลโค เอ (succinyl-CoA) ซักซิเนต (succinate) ฟูมาเรต (fumarate) มาเลต (malate) และออกซาโลอะซิเตด (oxaloacetate) พลังงานสุทธิเมื่อเกิดวัฏจักรเครปส์หนึ่งรอบ คือ 3 NADH, 1 FADH2 และ 1 GTP ซึ่ง GTP อาจสามารถถูกใช้สร้าง ATP ในภายหลังได้ ดังนั้น ผลผลิตสุทธิจากกลูโคสหนึ่งโมเลกุล (ไพรูเวต 2 โมเลกุล) คือ 6 NADH, 2 FADH2 และ 2 ATP จะได้น้ำ 12 โมเลกุล
ออกซิเดทีฟฟอสโฟรีเลชัน
ในยูคาริโอต ออกซิเดทีฟฟอสโฟรีเลชันเกิดขึ้นในคริสตีของไมโทคอนเดรีย ประกอบด้วย ลูกโซ่ของการขนส่งอิเล็กตรอน ซึ่งทำให้เกิดความแตกต่างของโปรตอน (ศักย์เคมิออสโมซิส) ข้ามเยื่อหุ้มชั้นในโดยการออกซิไดซ์ NADH ที่ได้จากวัฏจักรเครปส์ ATP จะถูกสังเคราะห์โดยเอนไซม์เอทีพีซินเทส (ATP synthase) เมื่อความแตกต่างทางเคมิออสโมซิสใช้ขับเคลื่อนฟอสโฟรีเลชันของ ADP ท้ายสุด อิเล็กตรอนจะถูกส่งต่อไปยังออกซิเจนที่หายใจรับเข้ามา และเมื่อรวมกับโปรตอนอีกสองตัว จะเกิดเป็นน้ำ
ตารางด้านล่างอธิบายปฏิกิริยาที่เกี่ยวข้องเมื่อกลูโคสหนึ่งโมเลกุลถูกออกซิไดซ์อย่างสมบูรณ์เป็นคาร์บอนไดออกไซด์ มีการสันนิษฐานว่า โคเอนไซม์ในรูปรีดิวซ์ทั้งหมดถูกออกซิไดซ์โดยลูกโซ่ของการขนส่งอิเล็กตรอนและถูกใช้ในออกซิเดทีฟฟอสโฟรีเลชัน
| ขั้น | ผลผลิตโคเอนไซม์ | ผลผลิต ATP | แหล่งของ ATP |
|---|---|---|---|
| ขั้นเตรียมของไกลโคไลสิส | -2 | ฟอสโฟรีเลชันของกลูโคสและฟรุกโทส 6-ฟอสเฟตใช้ 2 ATP จากไซโทพลาสซึม | |
| ขั้นได้พลังงานของไกลโคไลสิส | 4 | ฟอสโฟรีเลชันระดับซับสเตรต | |
| 2 NADH | 4–6 | ออกซิเดทีฟฟอสโฟรีเลชัน NADH แต่ละโมเลกุลสร้าง ATP สุทธิ 3 โมเลกุล จากการขนส่ง NADH ข้ามเยื่อหุ้มไมโทคอนเดรีย | |
| ออกซิเดทีฟดีคาร์บอกซิเลชันของไพรูเวต | 2 NADH | 6 | ออกซิเดทีฟฟอสโฟรีเลชัน |
| วัฏจักรเครปส์ | 2 | ฟอสโฟรีเลชันระดับซัสเตรต | |
| 6 NADH | 18 | ออกซิเดทีฟฟอสโฟรีเลชัน | |
| 2 FADH2 | 4 | ออกซิเดทีฟฟอสโฟรีเลชัน | |
| ผลผลิตทั้งหมด | 36–38 ATP | จากออกซิเดชันอย่างสมบูรณ์ของกลูโคสหนึ่งโมเลกุลเป็นคาร์บอนไดออกไซด์ และออกซิเดชันของโคเอนไซม์ในรูปรีดิวซ์ทั้งหมด | |
แม้ทางทฤษฎีจะระบุว่า กลูโคสหนึ่งโมเลกุลจะให้ผลผลิต ATP สุทธิสูงสุด 38 โมเลกุลจากการหายใจระดับบเซลล์ แต่สภาวะดังกล่าวโดยปกติไม่สามารถเกิดขึ้นได้จริง เนื่องจากการสูญเสีย เช่น ค่าการย้ายไพรูเวต ฟอสเฟตและ ADP เข้าสู่ไมโทคอนเดรีย ทั้งหมดต้องถูกลำเลียงแบบใช้พลังงานโดยใช้ตัวพาซึ่งใช้พลังงานที่เก็บสะสมไว้ในความแตกต่างทางไฟฟ้าเคมีของโปรตอน
ผลผลิตของขบวนการขนส่งเหล่านี้โดยใช้ความแตกต่างทางไฟฟ้าเคมีของโปรตอน คือ ต้องใช้มากกว่า 3 โปรตอนในการสร้าง 1 ATP จึงชัดเจนว่า เป็นการลดประสิทธิภาพของขบวนการทั้งหมดในทางทฤษฎี และผลผลิตสูงสุดที่น่าจะใกล้เคียงกับ 28-30 ATP มากกว่า ในทางปฏิบัติ ประสิทธิภาพอาจลดลงไปอีก เนื่องจากเยื่อหุ้มชั้นในของไมโทคอนเดรียนั้นรั่วต่อโปรตอนเล็กน้อย นอกจากนี้ยังมีปัจจัยอื่นอีก คือ โปรตีนอันคัปปลิง (uncoupling protein, UCP) ซึ่งมีอยู่ในบางเซลล์ เป็นช่องทางที่สามารถขนส่งโปรตอนได้ เมื่อโปรตีนนี้ทำงานในเยื่อหุ้มชั้นใน จะเป็นการลัดวงจรการจับคู่ระหว่างลูกโซ่ของการขนส่งอิเล็กตรอนกับการสังเคราะห์ ATP พลังงานศักย์จากความแกต่างของโปรตอนไม่ถกนำไปสร้าง ATP แต่จะให้ความร้อนแทน ซึ่งขบวนการดังกล่าวค่อนข้างมีความสำคัญในการก่อความร้อนของไขมันสีน้ำตาลในสัตว์แรกเกิดและจำศีล
การหมัก
เมื่อปราศจากออกซิเจน ไพรูเวตจะไม่เกิดเมแทบอลิซึมโดยการหายใจระดับเซลล์ แต่จะเกิดขบวนการการหมักแทน ไพรูเวตไม่ถูกขนส่งเข้าสู่ไมโทคอนเดรีย แต่จะยังอยู่ในไซโทพลาสซึม ที่ซึ่งมันจะถูกเปลี่ยนเป็นผลิตภัณฑ์ของเสียที่สามารถส่งออกนอกเซลล์ได้ ขบวนการดังกล่าวมีเพื่อออกซิไดซ์ NADH เป็น NAD+ จะได้นำกลับไปใช้ในไกลโคไลสิสได้ การหมักยังเป็นการป้องกันมิให้ NADH สะสมมากเกินไปในไซโทพลาสซึม ผลิตภัณฑ์ของเสียนี้แตกต่างกันไปในสิ่งมีชีวิตแต่ละชนิด ในกล้ามเนื้อลาย ผลิตภัณฑ์ของเสียที่ได้ คือ กรดแลกติก การหมักประเภทนี้ เรียกว่า การหมักกรดแลกติก ระหว่างไกลโคไลสิสแบบไม่ใช้ออกซิเจน NAD+ จะถูกสร้างใหม่เมื่อไฮโดรเจนหนึ่งคู่ไปรวมกับไพรูเวตเกิดเป็นแลกเตต การเกิดแลกเตตถูกเร่งปฏิกิริยาโดยแลกเตตดีไฮโดรจีเนส (lactate dehyrdrogenase) ในปฏิกิริยาผันกลับได้ แลกเตตยังสามารถใช้เป็นตัวตั้งต้นทางอ้อมสำหรับไกลโคเจนในตับ เมื่อกลับมาอยู่ในสภาวะที่มีออกซิเจน NAD+ ติดกับไฮโดรเจนจากแลกเตตเพื่อสร้าง ATP ในยีสต์ ผลิตภัณฑ์ของเสียที่ได้คือ เอทานอลและคาร์บอนไดออกไซด์ การหมักประเภทนี้ เรียกว่า การหมักแอลกอฮอล์หรือการหมักเอทานอล ATP ที่สร้างได้จากขบวนการนี้เป็นฟอสโฟรีเลชันระดับซับสเตรต ซึ่งไม่อาศัยออกซิเจน
การหมักเป็นขบวนการที่ด้อยประสิทธิภาพกว่าในการใช้พลังงานจากกลูโคส เพราะได้พลังงานออกมาเพียง 2 ATP ต่อกลูโคสเท่านั้น เมื่อเทียบกับ 38 ATP ที่ผลิตได้จากการหายใจแบบใช้ออกซิเจน ทั้งนี้เนื่องจากผลิตภัณฑ์ของเสียจากการหมักยังมีพลังงานสะสมอยู่มาก โปรคาริโอตที่ต้องคงอัตราการเจริญเติบโตที่รวดเร็วต่อไปเมื่อพวกมันย้ายจากสิ่งแวดล้อมที่มีออกซิเจนไปยังสิ่งแวดล้อมที่ขาดออกซิเจน พวกมันต้องเพิ่มอัตราปฏิกิริยาไกลโคไลสิส สำหรับสิ่งมีชีวิตหลายเซลล์ ระหว่างการหายใจกระชั้นในกิจกรรมที่ต้องใช้แรงมาก เซลล์กล้ามเนื้อจะใช้การหมักสร้าง ATP แทนการหายใจแบบใช้ออกซิเจนที่ช้ากว่า ดังนั้น เซลล์จึงอาจใช้การหมักแม้ก่อนระดับออกซิเจนจะหมดลง
การหายใจแบบไม่ใช้ออกซิเจน
จุลินทรีย์บางชนิดหายใจแบบไม่ใช้ออกซิเจน ซึ่งไม่มีออกซิเจน (การหายใจแบบใช้ออกซิเจน) หรือไพรูเวตหรืออนุพันธ์ของไพรูเวต (การหมัก) เป็นตัวรับอิเล็กตรอนตัวสุดท้าย แต่ใช้ตัวรับอิเล็กตรอนอนินทรีย์แทน เช่น ซัลเฟอร์