

ระบบการเห็น หรือ ระบบประสาทการมองเห็น หรือ ระบบสายตา(อังกฤษ: visual system) เป็นส่วนของระบบประสาทกลาง ซึ่งยังสิ่งมีชีวิตให้สามารถทำการรับรู้ด้วยการเห็น (visual perception) และทำให้เกิดการตอบสนองเนื่องด้วยข้อมูลทางตาแต่ไม่มีการรับรู้ด้วยการเห็นหลายอย่าง เป็นระบบที่สามารถตรวจจับและแปลผลข้อมูลแสงที่อยู่ในระหว่างความถี่ที่เห็นได้ เพื่อจะสร้างรูปแบบจำลองของสิ่งแวดล้อม ระบบการเห็นทำหน้าที่ที่ซับซ้อนหลายอย่างรวมทั้ง
- การรับแสงและการสร้างรูปแบบจำลองจากการเห็นด้วยตาแต่ละข้าง (monocular)
- การสร้างการรับรู้คือการเห็นด้วยตาทั้งสอง (binocular) จากข้อมูล 2-มิติ จากตาทั้งสองข้าง
- การรู้จำและการจำแนกประเภทของวัตถุที่เห็น
- การประเมินระยะทางที่จะไปถึงวัตถุและระยะทางระหว่างวัตถุ และ
- การนำการเคลื่อนไหวทางกายโดยสัมพันธ์กับวัตถุที่เห็น
กระบวนการประมวลข้อมูลทางตาตามแนวจิตวิทยา เรียกว่า การรับรู้ทางตา หรือ การรับรู้ด้วยการเห็น (visual perception) และความไม่สามารถรับรู้ด้วยการเห็นเรียกว่า ตาบอด กิจหน้าที่เกี่ยวข้องกับการเห็นแม้จะไม่ก่อให้เกิดการรับรู้ทางตา และเป็นอิสระจากการรับรู้ทางตา รวมกิจเช่น รีเฟร็กซ์รูม่านตาต่อแสง (pupillary light reflex) และ circadian photoentrainment
อารัมภบท
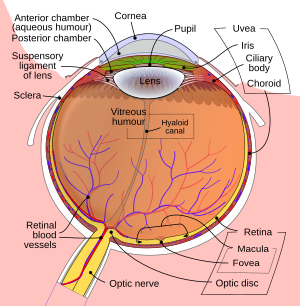
บทความนี้โดยมากพรรณนาถึงระบบการเห็นของสัตว์เลี้ยงลูกด้วยนม แม้ว่าสัตว์ซับซ้อนอย่างอื่นอาจจะมีระบบการเห็นที่คล้ายคลึงกัน ในสัตว์เลี้ยงลูกด้วยนม ระบบการเห็นแบ่งออกเป็น
- ตา โดยเฉพาะตัวเรตินา
- เส้นประสาทตา (Optic nerve)
- ส่วนไขว้ประสาทตา (Optic chiasma)
- ลำเส้นใยประสาทตา (Optic tract)
- นิวเคลียสงอคล้ายเข่าด้านข้าง (Lateral geniculate nucleus)
- ส่วนแผ่ประสาทตา (Optic radiation)
- คอร์เทกซ์สายตา (Visual cortex)
- คอร์เทกซ์สายตาสัมพันธ์ (Visual association cortex)
สัตว์สปีชีส์ต่าง ๆ สามารถเห็นส่วนต่าง ๆ ของสเปกตรัมแสงได้ไม่เหมือนกัน ตัวอย่างเช่น ผึ้งสามารถเห็นแสงอัลตราไวโอเลต ในขณะที่สัตว์ในวงศ์ย่อยงูหางกระดิ่งสามารถล่าสัตว์โดยใช้แอ่งรับรู้แสงอินฟราเรด ส่วนตาของปลากระโทงดาบสามารถสร้างความร้อนเพื่อตรวจจับเหยื่อให้ดียิ่งขึ้นในน้ำลึกมากกว่า 2,000 ฟุต
ประวัติ
ในครึ่งหลังแห่งคริสต์ศตวรรษที่ 19 มีการตั้งทฤษฎีต่าง ๆ เกี่ยวกับการทำงานในระบบประสาทเช่น neuron doctrine และ functional specialization ซึ่งเป็นทฤษฎีที่กำหนดว่า นิวรอนเป็นหน่วยพื้นฐานของระบบประสาท และว่า เขตต่าง ๆ ของสมองมีหน้าที่ความชำนาญที่ต่าง ๆ กัน ตามลำดับ ทฤษฎีเหล่านี้ได้กลายเป็นหลักของศาสตร์ใหม่คือประสาทวิทยาศาสตร์ และเป็นหลักสนับสนุนการทำความเข้าใจของระบบสายตา
ในปี ค.ศ. 1810 ฟรานซ์ โจเซฟ กอลล์ ได้เสนอความคิดเป็นครั้งแรกว่า เปลือกสมองแบ่งออกโดยกิจเป็นคอร์เทกซ์ส่วนต่าง ๆ ที่เดี๋ยวนี้รู้กันว่ามีหน้าที่เกี่ยวกับ การรับรู้สัมผัส (คือ คอร์เทกซ์รับรู้ความรู้สึกทางกาย) การสั่งการเคลื่อนไหว (คือ คอร์เทกซ์สั่งการ) และการเห็น (คือ คอร์เทกซ์การเห็น) และหลักฐานว่า สมองแบ่งออกโดยกิจเป็นเขตต่าง ๆ ก็เพิ่มพูนทวีคูณยิ่งขึ้นด้วยการค้นพบศูนย์ภาษาคือเขตโบรคา โดยพอล์ โบรคา ในปี ค.ศ. 1861 และการค้นพบคอร์เทกซ์สั่งการ (motor cortex) โดยกัสตาฟ ฟริตซ์ และเอดวา ฮิตซิก ในปี ค.ศ. 1871 ในปี ค.ศ. 1876 โดยใช้วิธีการทำลายส่วนในสมองและศึกษาผลที่ตามมา เดวิด เฟอร์ริเออร์ ได้เสนอว่า กิจหน้าที่ในการเห็นจำกัดอยู่ในสมองกลีบข้าง (parietal lobe) แต่ในปี ค.ศ. 1881 เฮอร์แมนน์ มังค์ เสนอว่า การเห็นนั้นอยู่ในสมองกลีบท้ายทอย (occipital lobe) ซึ่งเป็นข้อเสนอที่สมบูรณ์แม่นยำกว่าของเฟอร์ริเออร์ คือ สมองกลีบท้ายทอยเป็นส่วนที่เดี๋ยวนี้รู้กันว่า เป็นที่อยู่ของคอร์เทกซ์สายตาปฐมภูมิ (primary visual cortex)
ชีวภาพของระบบสายตา
ตา
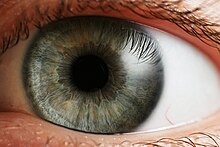
ตา คือ อวัยวะที่ทำหน้าที่รับแสง โดยสัตว์แต่ละชนิดจะมีอวัยวะรับแสงที่แตกต่างกัน ตาที่เรียบง่ายที่สุดจะไม่สามารถทำอะไรได้เลยเว้นแต่การรับรู้ว่าสิ่งแวดล้อมนั้นมืดหรือสว่างเพื่อให้สามารถปรับตัวให้เข้ากับการเปลี่ยนแปลงของสภาพแวดล้อมได้ เช่น กลางวันหรือกลางคืน เป็นต้น แต่จะไม่สามารถรับรู้ออกมาเป็นภาพได้ ตาที่ซับซ้อนกว่าจะมีรูปทรงและสีที่เป็นเอกลักษณ์ ในระบบตาที่ซับซ้อน ตาแต่ละดวงจะสามารถรับภาพที่มีบริเวณที่ซ้อนทับกันได้ เพื่อให้สมองสามารถรับรู้ถึงความลึก หรือ ความเป็นสามมิติของภาพ เช่น ระบบตาของมนุษย์ อย่างไรก็ตาม ตาของสัตว์บางชนิด เช่น กระต่ายและกิ้งก่า ได้ถูกออกแบบมาให้มีส่วนของภาพที่ซ้อนทับกันน้อยที่สุด แสงสว่างที่เข้าไปในตาจะมีการหักเหเมื่อผ่านกระจกตา และเมื่อแสงผ่านรูม่านตา (pupil) ซึ่งควบคุมโดยม่านตา (iris) ก็จะมีการหักเหเพิ่มขึ้นอีกโดยเลนส์ตา ทั้งกระจกตาและเลนส์รวมกันทำหน้าที่เป็นเลนส์แบบผสม เพื่อรวมแสงเป็นรูปกลับด้านให้ตกลงที่เรตินา
เรตินา

เรตินาประกอบด้วยเซลล์รับแสง (photoreceptor cell) เป็นจำนวนมากที่มีโมเลกุลโปรตีนที่เรียกว่า opsin ในมนุษย์ มี opsin 2 ชนิดที่เกี่ยวข้องกับการเห็นใต้อำนาจจิตใจ คือ rod opsins ในเซลล์รูปแท่ง (rod cell) และ cone opsins ในเซลล์รูปกรวย (cone cell) โปรตีนชนิดที่ 3 ที่เรียกว่า melanopsin อยู่ใน retinal ganglion cell (RGC) ซึ่งเป็นส่วนของกลไกการรักษาเวลาของร่างกาย และอาจจะไม่มีส่วนเกี่ยวข้องกับการเห็นใต้อำนาจจิตใจ เพราะว่า RGC ไม่ส่งสัญญาณไปยัง นิวเคลียสงอคล้ายเข่าด้านข้าง (lateral geniculate nucleus) แต่ส่งสัญญาณไปยัง pretectal area แทน
โปรตีน opsin หนึ่ง ๆ ทำหน้าที่รับโฟตอนหนึ่ง ๆ แล้วส่งสัญญาณต่อไปที่เซลล์ด้วยกรรมวิธี การถ่ายโอนสัญญาณ (signal transduction) มีผลเป็นการเพิ่มขั้ว (hyperpolarization) ในเซลล์รับแสง
เซลล์รูปแท่งและเซลล์รูปกรวยมีหน้าที่แตกต่างกัน เซลล์รูปแท่งปรากฏอย่างหนาแน่นในส่วนนอกของเรตินา ใช้ในการเห็นเมื่อมีแสงสลัว เซลล์รูปกรวยพบได้อย่างหนาแน่นที่ศูนย์กลางของเรตินา (คือที่ fovea) มีเซลล์รูปกรวย 3 ประเภทที่รับแสงโดยความถี่ที่ต่างกันโดยมีชื่อว่า short (สั้น) หรือ blue (น้ำเงิน), middle (กลาง) หรือ green (เขียว), และ long (ยาว) หรือ red (แดง) เซลล์รูปกรวยโดยหลักใช้เพื่อแยกแยะสีและลักษณะอื่น ๆ ของโลกทางตาในระดับแสงที่ปกติ
ในเรตินา เซลล์รับแสงเชื่อมไซแนปส์โดยตรงกับเซลล์สองขั้ว (bipolar cell) ซึ่งก็เชื่อมไซแนปส์กับ retinal ganglion cell (RGC) ในชั้นนอกสุดของเรตินา ซึ่งเป็นเซลล์ที่ส่งศักยะงานไปยังสมอง การประมวลผลทางตาเป็นจำนวนสำคัญ เกิดขึ้นเนื่องจากรูปแบบของการติดต่อสื่อสารระหว่างกันและกันของนิวรอนในเรตินา คือ แม้ว่า จะมีเซลล์รับแสงถึง 130 ล้านตัว แต่มีแอกซอนของ RGC เพียงแค่ 1.2 ล้านแอกซอนเท่านั้นที่ส่งข้อมูลจากเรตินาไปยังสมอง นั่นก็คือเรตินาไม่ได้ข้อมูลดิบจากเซลล์รับแสงไปยังสมองโดยตรง อีกอย่างหนึ่ง การประมวลผลในเรตินารวมถึงการจัดระเบียบลานสัญญาณของเซลล์สองขั้วและ ganglion cell เป็นแบบ center-surround และรวมทั้งการรวมสัญญาณ (convergence) และการแผ่สัญญาณ (divergence) จากเซลล์รับแสงไปยังเซลล์สองขั้ว นอกจากนั้นแล้ว นิวรอนอื่น ๆ ในเรตินาโดยเฉพาะเซลล์แนวนอน (horizontal cell) และเซลล์ขั้วสั้น (amacrine cell) ส่งสัญญาณแผ่ออกไปด้านข้าง คือไปยังนิวรอนติดกันที่อยู่ในชั้นเดียวกัน ซึ่งรวมกันมีผลเป็นลานสัญญาณที่มีความซับซ้อนมากขึ้น ที่มีลักษณะไม่แยกแยะสีแต่ไวต่อความเคลื่อนไหวก็ได้ มีลักษณะไวสีแต่ไม่แยกแยะความเคลื่อนไหวก็ได้
กลไกการสร้างสัญญาณการเห็น
เรตินาปรับตัวตามแสงโดยใช้เซลล์รูปแท่ง คือ ในที่มืด สารติดสียากคือ retinal ที่อยู่ในโปรตีน opsin จะมีรูปคดเรียกว่า cis-retinal เมื่อเกิดมีแสงขึ้น retinal จะเปลี่ยนรูปไปเป็นแบบตรงเรียกว่า trans-retinal และจะแยกตัวออกจาก opsin ปรากฏการณ์นี้เรียกว่า bleaching (การฟอกขาว) เพราะว่า rhodopsin ที่ปราศจาก retinal เปลี่ยนสีจากสีม่วงไปเป็นไม่มีสีในที่ที่มีแสง ในระดับความมืดที่เป็นปทัฏฐาน rhodopsin ไม่มีการดูดแสง และจะหลั่งกลูตาเมตซึ่งมีฤทธิ์ยับยั้งเซลล์สองขั้ว การยับยั้งเซลล์สองขั้ว ก็จะเข้าไปยับยั้งการหลั่งสารสื่อประสาทจากเซลล์สองขั้วไปยัง RGC แต่เมื่อเกิดมีแสงขึ้น การหลั่งกลูตาเมตจากเซลล์รูปแท่งก็จะหยุดลง เซลล์สองขั้วก็จะหลั่งสารสื่อประสาทไปกระตุ้น RGC และดังนั้น กระบวนการตรวจจับสิ่งที่เห็นก็ได้เริ่มเกิดขึ้น ผลสุดท้ายของการประมวลผลอย่างนี้ก็คือ มีกลุ่มของ RGC 5 กลุ่มที่ส่งข้อมูลทางตา (ทั้งที่ทำให้เกิดการเห็นโดยที่สุด ทั้งที่ไม่ทำให้เกิดการเห็น) ไปยังสมอง กลุ่มทั้ง 5 คือ
- เซลล์ M มีลานสัญญาณแบบ center-surround ขนาดใหญ่ที่มีความไวต่อความลึก (คือระยะทางจากวัตถุที่เห็น) ไม่แยกแยะสี และสามารถปรับตัวตามตัวกระตุ้นได้อย่างรวดเร็ว
- เซลล์ P มีลานสัญญาณแบบ center-surround ขนาดที่เล็กกว่าที่ไวต่อทั้งสีและทั้งรูปร่าง
- เซลล์ K มีลานสัญญาณแบบ center-only ขนาดใหญ่มากที่ไวต่อสีและไม่แยกแยะรูปร่างหรือความลึก
- ganglion cell ประเภทไวแสง (photosensitive ganglion cell)
- เซลล์อีกกลุ่มหนึ่งที่ส่งสัญญาณใช้ในการเคลื่อนไหวตา
งานวิจัยปี ค.ศ. 2006 ที่มหาวิทยาลัยเพนซิลเวเนีย ประเทศสหรัฐอเมริกา ประเมินอัตราการส่งถ่ายข้อมูล (bandwidth) ของเรตินาในมนุษย์ว่าอยู่ที่ประมาณ 8,960 กิโลบิตต่อวินาที เปรียบเทียบกับในหนูตะเภาที่ประมาณ 875 กิโลบิตต่อวินาที
ในปี ค.ศ. 2007 ไซดีและคณะร่วมงานทำงานวิจัยในคนไข้ที่ปราศจากทั้งเซลล์รูปแท่งและเซลล์รูปกรวย และพบว่า ganglion cell ไวแสงที่พึ่งค้นพบใหม่ ๆ มีบทบาทในการรับรู้ทางตาทั้งที่อยู่ใต้อำนาจจิตใจและทั้งที่อยู่เหนืออำนาจจิตใจ เซลล์เหล่านั้นไวแสงที่สุดต่อแสงมีความยาวคลื่น 481 นาโนเมตร ผลงานวิจัยนี้แสดงว่า มีวิถีประสาทสองทางสำหรับข้อมูลการเห็นจากเรตินา ทางหนึ่งมีเซลล์รับแสงคือเซลล์รูปแท่งและเซลล์รูปกรวยเป็นฐาน และอีกทางหนึ่งมี ganglion cell ไวแสงที่พึ่งค้นพบใหม่ ทำหน้าที่เป็นเครื่องตรวจจับความสว่างอย่างคร่าว ๆ ทางตา
กระบวนการเคมีแสง
ในระบบการเห็น สาร retinal หรือที่มีชื่อในวงการว่า retinene1 หรือ retinaldehyde เป็นโมเลกุลไวแสงพบในเซลล์รูปแท่งและเซลล์รูปกรวยในเรตินา retinal เป็นโครงสร้างพื้นฐานที่มีหน้าที่ถ่ายโอนแสงไปเป็นสัญญาณประสาททางตา ซึ่งก็คือ พลังประสาทที่ผ่านระบบประสาทตาไปยังสมอง ในที่มีแสง โมเลกุลของ retinal เปลี่ยนรูปร่างไป ซึ่งทำให้เกิดการส่งพลังประสาทเป็นผลที่สุด
ใยประสาทตาสู่ทาลามัส
เส้นประสาทตา
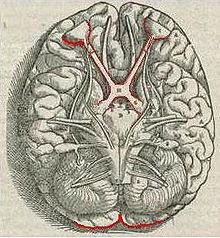
ตาส่งข้อมูลการเห็นไปยังสมองผ่านเส้นประสาทตา (Optic nerve) คือ กลุ่มต่าง ๆ ของ ganglion cell ในเรตินาส่งข้อมูลไปยังสมองผ่านเส้นประสาทตา ประมาณ 90% ของแอกซอนในเส้นประสาทตาไปสุดที่นิวเคลียสงอคล้ายเข่าด้านข้าง (lateral geniculate nucleus) ในทาลามัส แอกซอนเหล่านี้มีต้นกำเนิดจาก ganglion cell ประเภท M, P และ K ดังที่กล่าวมาแล้ว การประมวลผลแบบขนาน (คือหลาย ๆ แขนงหลาย ๆ ทางพร้อมกัน) แบบนี้ มีความสำคัญในการสร้างแบบจำลองของโลกของการเห็น ข้อมูลแต่ละอย่างจะไปตามทางประสาทของตน ๆ ซึ่งนำไปสู่การรับรู้ (perception) โดยที่สุด ส่วนแอกซอนอีกกลุ่มหนึ่งส่งข้อมูลไปยัง superior colliculus ในสมองส่วนกลาง (midbrain) ซึ่งมีบทบาทในการสั่งการเคลื่อนไหวตาประเภท saccade และในปฏิกิริยาด้วยการเคลื่อนไหวอย่างอื่น ๆ อีกด้วย
ganglion cell ไวแสงกลุ่มหนึ่ง ซึ่งมีสารไวแสง melanopsin ส่งข้อมูลทางตาไปทางประสาท retinohypothalamic tract ไปยัง pretectum (มีบทบาทเกี่ยวกับรีเฟล็กซ์รูม่านตา), ไปยังโครงสร้างอื่น ๆ ที่มีบทบาทในการควบคุมจังหวะรอบวัน (circadian rhythm) และการนอนหลับ เช่น suprachiasmatic nucleus (มีบทบาทเป็นนาฬิกาทางชีวภาพ), และไปยัง ventrolateral preoptic nucleus ซึ่งมีบทบาทในการควบคุมการนอนหลับ บทบาทที่พึ่งค้นพบเร็ว ๆ นี้ของ ganglion cell ไวแสงก็คือ เป็นตัวสื่อการเห็นทั้งภายใต้อำนาจจิตใจและทั้งเหนืออำนาจจิตใจ คือทำหน้าที่เป็นเครื่องตรวจจับความสว่างอย่างคร่าว ๆ ทางตา
ส่วนไขว้ประสาทตา
เส้นประสาทตาจากตาทั้งสองเดินทางมาบรรจบและข้ามแทยงกันที่ส่วนไขว้ประสาทตา (Optic chiasma) ที่ฐานของไฮโปทาลามัสในสมอง (ดูรูปข้างบน) ที่ส่วนไขว้นี้ ข้อมูลจากทั้งสองตามารวมกันแล้วก็แยกกันไปตามลานสายตา (visual field) คือ ข้อมูลลานสายตาด้านซ้ายส่งไปยังสมองซีกขวา และข้อมูลลานสายตาด้านขวาส่งไปยังสมองซีกซ้าย เพื่อการประมวลผลต่อไป นั่นก็คือ คอร์เทกซ์สายตาปฐมภูมิซีกขวา จัดการข้อมูลจากลานสายตาด้านซ้าย แม้ว่าข้อมูลลานสายตาจะมาจากตาทั้งสองข้าง และโดยนัยเดียวกัน คอร์เทกซ์สายตาปฐมภูมิอีกซีกหนึ่งก็เช่นกัน แต่ว่า ก็ยังมีเขตเล็ก ๆ ท่ามกลางลานสายตาที่มีการประมวลผลซ้ำซ้อนกันโดยซีกสมองทั้งสองข้าง
ลำเส้นใยประสาทตา
ข้อมูลจากลานสายตาด้านขวา (ซึ่งมาถึงด้านซ้ายของสมองแล้ว) เดินทางไปในลำเส้นใยประสาทตา (optic tract) ด้านซ้าย และข้อมูลจากลานสายตาด้านซ้าย ก็เดินทางไปในลำเส้นใยประสาทตา ด้านขวา (ดูรูปข้างบน) ลำเส้นใยประสาทตาแต่ละเส้นไปสุดที่นิวเคลียสงอคล้ายเข่าด้านข้าง (lateral geniculate nucleus) ในทาลามัส
นิวเคลียสงอคล้ายเข่าด้านข้าง
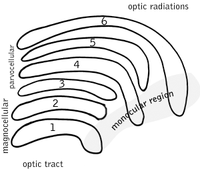
นิวเคลียสงอคล้ายเข่าด้านข้าง (lateral geniculate nucleus ตัวย่อ LGN) เป็นนิวเคลียสถ่ายทอดสัญญาณความรู้สึก (จากตาไปยังคอร์เทกซ์สายตาปฐมภูมิ) ในทาลามัส LGN ประกอบดัวยชั้น 6 ชั้นในมนุษย์และในไพรเมตบางประเภทเริ่มต้นแต่ไพรเมตที่มี Parvorder เป็น Catarrhini รวมทั้งวงศ์ลิงโลกเก่าและเอป ชั้น 1, 4 และ 6 เป็นชั้นที่รับข้อมูลมาจากใยประสาทด้านตรงข้ามกายจากเรตินาใกล้จมูก (หรือจากลานสายตาใกล้ขมับ) ส่วนชั้น 2, 3 และ 5 รับข้อมูลจากใยประสาทจากเรตินาด้านขมับที่อยู่ในกายด้านเดียวกัน (หรือจากลานสายตาใกล้จมูก)
ชั้นที่ 1 ของ LGN ประกอบด้วยเซลล์ M ซึ่งรับข้อมูลมาจากเซลล์ M (magnocellular) ของเส้นประสาทตาจากตาด้านตรงข้าม และมีข้อมูลเกี่ยวกับความลึกและการเคลื่อนไหว ชั้นที่ 4 และ 6 ของ LGN ก็เชื่อมต่อกับตาด้านตรงข้ามเช่นกัน แต่ว่าเชื่อมกับเซลล์ P ของเส้นประสาทตาที่มีข้อมูลเกี่ยวกับสีและเส้นขอบ (ของสิ่งที่เห็น) เปรียบเทียบกับ ชั้น 2, 3 และ 5 ของ LGN ที่เชื่อมกับเซลล์ M และ P (parvocellular) ของเส้นประสาทตาจากกายด้านเดียวกัน เมื่อแผ่ออก ชั้นทั้งหกของ LGN มีขนาดเท่ากันกับบัตรเครดิตแต่มีความหนาประมาณ 3 เท่าของบัตร เมื่อม้วนเข้า LGN มีขนาดและรูปร่างเป็นไข่นก 2 ใบเล็ก ๆ และในระหว่างชั้นทั้ง 6 มีชั้นของเซลล์เล็ก ๆ ที่รับข้อมูลมาจากเซลล์ K ซึ่งมีข้อมูลเกี่ยวกับสีจากเรตินา
แม้ว่า นิวรอนของ LGN จะถ่ายทอดรูปทางตาต่อไปยังคอร์เทกซ์สายตาปฐมภูมิ (หรือ V1) ซึ่งอยู่ในสมองกลีบท้ายทอย ใน calcarine sulcus และบริเวณรอบ ๆ นั้น แต่ว่า LGN อาจไม่ใช่เป็นเพียงแค่สถานีถ่ายทอดสัญญาณไปยัง V1 เท่านั้น แต่ตัวเองก็ยังอาจเป็นศูนย์การประมวลผลด้วย คือ LGN รับข้อมูลป้อนกลับจาก V1 เป็นจำนวนประมาณ 10 เท่าของข้อมูลที่ส่งไป และถึงแม้ว่า ยังไม่มีความชัดเจนว่า การประมวลผลนี้ให้ผลเป็นอะไร แต่นักวิจัยบางพวกเชื่อว่าการเชื่อมต่อเป็นวงจรอย่างนี้ ช่วยการพยากรณ์ความมีอยู่ของตัวกระตุ้น เป็นกระบวนการที่เรียกว่า การเข้ารหัสตัวกระตุ้นด้วยการพยากรณ์ (predictive coding) และงานวิจัยปี ค.ศ. 2000 แสดงว่า การหยุดการทำงานของคอร์เทกซ์สายตาปฐมภูมิของแมวโดยใช้ความเย็น มีผลต่อเส้นโค้งการตอบสนองของความเปรียบต่าง (contrast response curve) ของ LGN
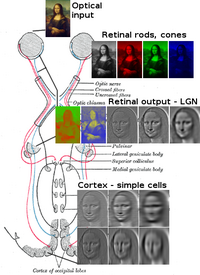
ส่วนแผ่ประสาทตา
ส่วนแผ่ประสาทตา (Optic radiation) ซึ่งมีอยู่ในซีกสมองทั้งสองข้าง ส่งข้อมูลไปจากนิวเคลียสงอคล้ายเข่าด้านข้าง (lateral geniculate nucleus ตัวย่อ LGN) ที่อยู่ในทาลามัส ไปยังคอร์เทกซ์สายตาปฐมภูมิ (หรือ V1) คือ นิวรอนชั้น P ของ LGN ส่งข้อมูลไปยังชั้น 4Cβ ของ V1, ส่วนนิวรอนชั้น M ส่งข้อมูลไปยังชั้น 4Cα และ 6, และนิวรอนชั้น K ส่งข้อมูลไปให้กับกลุ่มนิวรอนที่เรียกว่า blobs ในชั้น 2 และ 3
ในแต่ละจุดของลานสายตา จะมีใยประสาทตาในลำเส้นใยประสาทตา และจะมีนิวรอนใน V1 ที่สัมพันธ์กับจุดนั้น แต่ว่าหลังจากนั้น ความสัมพันธ์อย่างตรงไปตรงมาแบบนี้จะเริ่มหายไป เนื่องจากว่า มีการเชื่อมต่อกันอย่างข้ามไปข้ามมาระหว่างนิวรอนในคอร์เทกซ์สายตา
คอร์เทกซ์สายตา
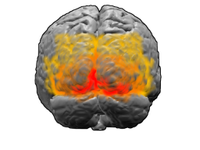
คอร์เทกซ์สายตาเป็นระบบสมองที่ใหญ่ที่สุดในมนุษย์ และมีหน้าที่ประมวลผลเกี่ยวกับสิ่งที่เห็นทางตา อยู่ด้านท้ายของสมอง (ซึ่งได้รับการเน้นในรูปข้างบน) เหนือซีรีเบลลัม เขตในคอร์เทกซ์ที่รับการเชื่อมต่อโดยตรงจาก LGN เรียกว่าคอร์เทกซ์สายตาปฐมภูมิ (หรือเรียกว่า V1 หรือคอร์เทกซ์ลาย) ข้อมูลทางตาผ่าน V1 เข้าไปสู่เขตสายตาต่าง ๆ ตามลำดับ ซึ่งก็คือเขต V2, V3, V4 และ V5 (หรือที่เรียกว่า MT) แต่ว่า การเชื่อมต่อกันระหว่างเขตเหล่านี้จะต่าง ๆ กันไปในสัตว์ขึ้นอยู่กับสปีชีส์ เขตสายตาทุติยภูมิเหล่านี้ ซึ่งเรียกรวมกันว่า เขตคอร์เทกซ์สายตานอกคอร์เทกซ์ลาย (extrastriate visual cortex) ทำหน้าที่ประมวลผลจากข้อมูลพื้นฐานทางตามากมาย นิวรอนใน V1 และ V2 ตอบสนองโดยเฉพาะต่อเส้นที่มีทิศทางโดยเฉพาะ (เช่นเส้นตั้ง เส้นนอน) หรือส่วนประกอบที่มีเส้นเหล่านั้น ความเป็นไปอย่างนี้ เชื่อกันว่า เป็นเพื่อสนับสนุนการตรวจจับขอบและมุมของวัตถุที่เห็น โดยนัยเช่นเดียวกัน ข้อมูลพื้นฐานเกี่ยวกับสีและความเคลื่อนไหวก็ได้รับการประมวลผลใน V1
คอร์เทกซ์สายตาสัมพันธ์
ในขณะที่ข้อมูลทางตาเดินทางไปในเขตต่าง ๆ ของคอร์เทกซ์สายตา ระดับความซับซ้อนของข้อมูลต่าง ๆ ที่นิวรอนรองรับ (คือเป็นตัวแทน) ก็เพิ่มขึ้นเรื่อย ๆ เปรียบเทียบกับนิวรอนใน V1 ที่อาจจะตอบสนองโดยเฉพาะต่อเส้นที่มีทิศทางโดยเฉพาะในลานสายตาส่วนเฉพาะ (ที่เป็นส่วนของวัตถุที่เห็น) นิวรอนด้านข้างของสมองกลีบท้ายทอยอาจจะตอบสนองโดยเฉพาะต่อวัตถุที่เห็นทั้งหมด (เช่นรูปวาด) และนิวรอนในคอร์เทกซ์สายตาสัมพันธ์อาจจะตอบสนองโดยเฉพาะต่อใบหน้ามนุษย์ หรือต่อวัตถุใดวัตถุหนึ่ง
เมื่อความซับซ้อนของข้อมูลที่รองรับโดยนิวรอนเพิ่มขึ้นเรื่อย ๆ อย่างนี้ ก็อาจจะเริ่มมีการประมวลผลแบบเฉพาะหน้าที่ในระบบนิวรอนที่แบ่งออกเป็นทางประสาท 2 ทาง คือทางสัญญาณด้านหลัง (dorsal stream) และทางสัญญาณด้านล่าง (ventral stream) ซึ่งเป็นทางสัญญาณที่เสนอไว้ในสมมุติฐานทางสัญญาณสองทาง (Two-streams hypothesis) ซึ่งเป็นสมมุติฐานที่เสนอเป็นครั้งแรกโดย เลสลี อังเกอร์เลเดอร์ และมอร์ติเมอร์ มิชกิน ในปี ค.ศ. 1982 ทางสัญญาณด้านหลัง ที่นิยมระบุว่า "เป็นทางสัญญาณบอกว่า ที่ไหน" มีบทบาทในการใส่ใจเกี่ยวกับปริภูมิ และเชื่อมต่อกับเขตสมองอื่น ๆ ที่สั่งการการเคลื่อนไหวตาและมือ นอกจากนั้นในเร็ว ๆ นี้ ทางสัญญาณนี้ยังได้รับชื่อว่า "ทางสัญญาณบอกว่า อย่างไร" เพื่อเน้นบทบาทในพฤติกรรมที่นำการเคลื่อนไหวร่างกายไปในส่วนต่าง ๆ ในปริภูมิ เปรียบเทียบกับทางสัญญาณด้านล่าง ที่นิยมระบุว่า "เป็นทางสัญญาณบอกว่า อะไร" ที่มีบทบาทในการรู้จำ ในการระบุ และในการจัดประเภทของสิ่งที่เห็น
อย่างไรก็ดี ก็ยังมีการอภิปรายกันถึงระดับของการทำงานเฉพาะหน้าที่ในทางสัญญาณทั้งสองนี้ เนื่องจากว่า ทางสัญญาณทั้งสองนี้มีการเชื่อมต่อไปหากันและกันอย่างหนาแน่น
การจัดประเภทของวัตถุที่เห็น จากงานวิจัยใช้ fMRI
อเล็กซานเดอร์ ฮูธ และคณะ ที่มหาวิทยาลัยแคลิฟอร์เนียเบิร์กลีย์ ได้ทำงานวิจัยในมนุษย์ 5 คนที่ดูคลิปภาพยนตร์เป็นระยะเวลา 2 ช.ม. ในขณะที่มีการสร้างภาพสมองโดยเทคนิค fMRI ไปในขณะเดียวกัน การสร้างภาพสมองแต่ละครั้งบันทึกระดับการเดินโลหิตในส่วนต่าง ๆ ของสมองเป็นพัน ๆ ส่วน และเมื่อแปลงผลข้อมูลที่ได้โดยใช้วิธี Principal components analysis ก็ปรากฏว่าคอร์เทกซ์มีตำแหน่งกว่า 30,000 ตำแหน่งที่รองรับการจัดประเภทของสิ่งที่เห็นมากกว่า 1,700 ประเภท ฮูธและคณะได้ค้นพบแผนที่ (แสดงประเภทของสิ่งที่เห็น) ที่คาบเกี่ยวกัน ที่เป็นระเบียบอย่างยิ่ง ในพื้นที่มากกว่า 20% ในคอร์เทกซ์
ระบบการเห็นและการทรงตัว
พร้อมกับระบบการรับรู้อากัปกิริยา (proprioception) และ vestibular system ระบบการเห็นมีบทบาทสำคัญในการที่บุคคลสามารถควบคุมการทรงตัวและรักษาอากัปกิริยาที่ตั้งตรงไว้ได้ เมื่อแยกระบบสามอย่างเหล่านี้เป็นส่วน ๆ เพื่อที่จะทดสอบหน้าที่และอิทธิพลในการทรงตัว พบว่า การเห็นเป็นองค์ประกอบที่สำคัญที่สุดในการทรงตัว คือ สำคัญกว่าระบบทั้ง 2 ที่เหลือ ความชัดเจนที่บุคคลหนึ่งสามารถเห็นสิ่งที่อยู่แวดล้อม ขนาดของลานสายตา (visual field) ความอ่อนไหวต่อแสงบาดตา และความสามารถในการรับรู้ความลึก (คือระยะทางจากตัว) มีบทบาทสำคัญในการให้ข้อมูลป้อนกลับ (feedback) แก่สมอง เกี่ยวกับการเคลื่อนไหวของกายโดยสัมพันธ์กับสิ่งแวดล้อม อะไรก็ตามที่มีอิทธิพลต่อตัวแปรเหล่านี้ สามารถทำให้เกิดผลลบแก่การทรงตัวและการทรงอากัปกิริยา ผลลบเช่นนี้ปรากฏในงานวิจัยกับผู้ใหญ่วัยชรา เมื่อเปรียบเทียบกับชนกลุ่มควบคุมที่มีวัยเยาว์ ในคนไข้โรคต้อหินเปรียบเทียบกับชนกลุ่มทดลองมีวัยเดียวกัน ในคนไข้ต้อกระจกเทียบกันช่วงก่อนและหลังศัลยกรรมเพื่อรักษาต้อ และแม้แต่ในเหตุการณ์ปกติธรรมดาเช่นการใส่แว่นนิรภัย การเห็นด้วยตาข้างเดียว (monocular vision) มีผลลบต่อการทรงตัว ดังปรากฏในงานวิจัยเกี่ยวกับคนไข้ต้อหินและต้อกระจกดังที่กล่าวไปแล้ว และในเด็กและผู้ใหญ่ที่ปกติด้วย
ตามงานวิจัยในปี ค.ศ. 2010 ของโปลล็อกและคณะ โรคหลอดเลือดสมองเป็นเหตุหลักในความเสื่อมทางการเห็น และที่เกิดบ่อยที่สุดคือการสูญเสียส่วนในลานสายตา (visual field loss) เช่นการสูญเสียการเห็นประเภท homonymous hemianopsia แต่ว่า ยังเป็นที่น่าเสียดายว่า หลักฐานแสดงประสิทธิภาพของวิธีรักษาที่มีค่าใช้จ่ายที่เหมาะสม ที่ได้สร้างขึ้นเพื่อแก้ไขปัญหาลานสายตาเหล่านี้ ยังไม่มีความชัดเจน
ดูเพิ่ม
- ภาวะไม่รู้ความเคลื่อนไหว (Akinetopsia)
- ภาวะเสียการระลึกรู้ทางตาแบบวิสัญชาน (Apperceptive visual agnosia)
- ภาวะเสียการระลึกรู้ทางตาแบบสัมพันธ์ (Associative visual agnosia)
- สายตาเอียง (Astigmatism)
- ตาบอดสี (Color blindness)
- ภาวะไม่รู้ใบหน้า (Prosopagnosia)
- ภาวะเสียการระลึกรู้ทางตา (Visual agnosia)
อ่านเพิ่ม
- Davison JA, Patel AS, Cunha JP, Schwiegerling J, Muftuoglu O (July 2011). "Recent studies provide an updated clinical perspective on blue light-filtering IOLs". Graefes Arch. Clin. Exp. Ophthalmol. 249 (7): 957–68. doi:10.1007/s00417-011-1697-6. PMC 3124647. PMID 21584764.
- Hatori M, Panda S (October 2010). "The emerging roles of melanopsin in behavioral adaptation to light". Trends Mol Med. 16 (10): 435–46. doi:10.1016/j.molmed.2010.07.005. PMC 2952704. PMID 20810319.
- Heiting, G., (2011). Your infant’s vision Development. Retrieved February 27, 2012 from http://www.allaboutvision.com/parents/infants.htm
- Hubel, David H. (1995). Eye, brain, and vision. New York: Scientific American Library. ISBN 978-0-7167-6009-2. OCLC 32806252.
- Kolb B, Whishaw I (2012). Introduction to Brain and Behaviour Fourth Edition. New York: Worth Publishers. ISBN 978-1-4292-4228-8. OCLC 918592547.
- Marr, David; Ullman, Shimon; Poggio, Tomaso (2010). Vision: A Computational Investigation into the Human Representation and Processing of Visual Information. Cambridge, Mass: The MIT Press. ISBN 978-0-262-51462-0. OCLC 472791457.
- Rodiek, R.W. (1988). "The Primate Retina". Comparative Primate Biology. Neurosciences. New York: A.R. Liss. 4.. (H.D. Steklis and J. Erwin, editors.) pp. 203–278.
-
Schmolesky, Matthew (1995). "The Primary Visual Cortex". PMID 21413385. คลังข้อมูลเก่าเก็บจากแหล่งเดิมเมื่อ 2004-12-29. สืบค้นเมื่อ 2005-01-01.
{{cite journal}}: Cite journal ต้องการ|journal=(help). - The Aging Eye; See into Your future. (2009). Retrieved February 27, 2012 from https://web.archive.org/web/20111117045917/http://www.realage.com/check-your-health/eye-health/aging-eye
- Tovée, Martin J. (2008). An introduction to the visual system. Cambridge, UK: Cambridge University Press. ISBN 978-0-521-88319-1. OCLC 185026571.
-
Vesalius, Andreas (1543). "De Humani Corporis Fabrica (On the Workings of the Human Body)".
{{cite journal}}: Cite journal ต้องการ|journal=(help) - Wiesel, Torsten; Hubel, David H. (1963). "The effects of visual deprivation on the morphology and physiology of cell's lateral geniculate body". Journal of Neurophysiology. 26 (6): 978–993. doi:10.1152/jn.1963.26.6.978. PMID 14084170. S2CID 16117515.