
| เซอร์ปิน (สารยับยั้งเอนไซม์ซีรีนโปรตีเอส) | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
 ภาพแสดงโปรตีนเซอร์ปิน (สีขาว) ซึ่งมีวงศูนย์ปฏิกิริยา (reactive centre loop; สีน้ำเงิน) จับอยู่กับเอนไซม์โปรตีเอส (สีเทา). เมื่อเอนไซม์โปรตีเอสพยายามที่จะเข้าเร่งปฏิกิริยา เอนไซม์ดังกล่าวจะถูกยับยั้งการทำงานโดยเซอร์ปิน ซึ่งไม่สามารถผันกลับได้ (PDB 1K9O)
| |||||||||||
| ข้อมูลจำเพาะ | |||||||||||
| สัญลักษณ์ | Serpin, SERPIN (root symbol of family) | ||||||||||
| Pfam | PF00079 | ||||||||||
| InterPro | IPR000215 | ||||||||||
| PROSITE | PDOC00256 | ||||||||||
| SCOPe | 1hle / SUPFAM | ||||||||||
| CDD | cd00172 | ||||||||||
| |||||||||||
เซอร์ปิน (อังกฤษ: Serpin) เป็นมหาวงศ์ของกลุ่มโปรตีนที่มีโครงสร้างคล้ายคลึงกัน ถูกค้นพบเป็นครั้งแรกจากการที่กลุ่มโปรตีนดังกล่าวสามารถออกฤทธิ์เป็นสารยับยั้งเอนไซม์โปรตีเอส โดยกลุ่มโปรตีนเซอร์ปินนี้พบได้ในสิ่งมีชีวิตในทุกอาณาจักร ทั้งนี้ ชื่อ เซอร์ปิน นั้นถูกตั้งมาจากการที่เซอร์ปินชนิดแรกที่ถูกค้นพบออกฤทธิ์ต้านซีรีนโปรตีเอสที่มีโครงสร้างคล้ายไคโมทริปซิน (serine protease inhibitors) โปรตีนในมหาวงศ์นี้มีกลไกการออกฤทธิ์ที่โดดเด่นซึ่งไม่ค่อยพบเห็นได้ทั่วไปนัก กล่าวคือ เมื่อเซอร์ปินเข้าจับกับเอนไซม์เป้าหมาย การทำงานของเอนไซม์นั้นจะถูกยับยั้งแบบไม่สามารถผันกลับได้ (irreversibly inhibition) โดยทำให้เกิดการเปลี่ยนโครงรูปขนาดใหญ่บนตำแหน่งกัมมันต์ของเอนไซม์ที่ถูกจับ ซึ่งแตกต่างจากกลไกการยับยั้งแข่งขันทั่ว ๆ ไปของสารยับยั้งเอนไซม์โปรตีเอสที่มักเข้าจับและขัดขวางการเข้าถึงตำแหน่งกัมมันต์บนเอนไซม์
การยับยั้งเอนไซม์โปรตีเอสของเซอร์ปินนั้นส่งผลต่อกระบวนการทางชีวภาพของสิ่งมีชีวิตหลายประการ ซึ่งรวมไปถึงการจับลิ่มของเลือดและการอักเสบ จึงทำให้โปรตีนเหล่านี้ได้รับความสนใจเป็นอย่างมากในการศึกษาวิจัยทางการแพทย์ นอกจากนี้ ด้วยการออกฤทธิ์ด้วยการเปลี่ยนโครงรูปที่เป็นเอกลักษณ์ ยังทำให้กลุ่มโปรตีนนี้เป็นที่สนใจในกลุ่มการวิจัยด้านชีววิทยาเชิงโครงสร้างและการม้วนพับโปรตีนอีกด้วย อย่างไรก็ตาม ถึงแม้การออกฤทธิ์เพื่อให้เกิดการเปลี่ยนโครงรูปของเซอร์ปีนจะมีข้อดีหลายประการ แต่ก็มีข้อด้อยเช่นเดียวกัน โดยเซอร์ปีนอาจมีความเสี่ยงต่อการกลายพันธุ์เพิ่มขึ้น ซึ่งจะส่งผลให้เกิดโรคจากความผิดปกติของเซอร์ปิน (serpinopathies) อันเป็นผลต่อเนื่องมาจากการม้วนพับโปรตีนที่ผิดปกติ และการสร้างพอลิเมอร์สายยาวที่ไม่สามารถทำงานได้ การเกิดพอลิเมอไรเซชันของเซอร์ปิน นอกจากจะทำให้สารยับยั้งที่ออกฤทธิ์ (active inhibitor) มีปริมาณลดน้อยลงแล้ว ยังทำให้เกิดการตายของเซลล์และอวัยวะล้มเหลวจากการสะสมของพอลิเมอร์เหล่านั้นด้วย
ถึงแม้เซอร์ปินส่วนใหญ่จะควบคุมกระบวนการการย่อยสลายโปรตีน (proteolytic cascades) ยังมีโปรตีนบางชนิดที่มีโครงสร้างคล้ายเซอร์ปินที่ไม่ได้ออกฤทธิ์เป็นสารยับยั้งเอนไซม์โปรตีเอส แต่ทำหน้าที่อื่นที่แตกต่างออกไป ตัอย่างเช่น เป็นโปรตีนเก็บสะสม (storage protein) เช่นที่พบในไข่ขาว เรียกว่าโอแวลบูมิน (ovalbumin), เป็นโปรตีนขนส่ง (carriage proteins) สำหรับการขนส่งฮอร์โมนบางชนิดไปยังตำแหน่งออกฤทธิ์ (thyroxine-binding globulin, cortisol-binding globulin หรือ transcortin เป็นอาทิ) และทำหน้าที่เป็นแชพเพอโรน (เช่น HSP47) โดยมีการใช้คำว่า เซอร์ปิน เพื่ออธิบายถึงกลุ่มโปรตีนข้างต้นอยู่เช่นกัน แม้จะไม่ได้ทำหน้าที่ในการยับยั้งเอนไซม์โปรตีเอสก็ตาม
ประวัติ
การค้นพบกระบวนการยับยั้งเอนไซม์โปรตีเอสถูกรายงานเป็นครั้งแรกในคริสตวรรษที่ 18 แต่นับจากนั้นก็ไม่พบรายงานการศึกษาวิจัยที่เกี่ยวข้องกับกระบวนการดังกล่าวอีกเลย จนกระทั่งใน ปี ค.ศ. 1955 ได้มีการคัดแยกสารเซอร์ปิน 2 ชนิดได้สำเร็จ ได้แก่ แอนติทรอมบิน และแอลฟา-1 แอนตีทริปซิน ทั้งนี้ การศึกษาวิจัยเกี่ยวกับเซอร์ปินในช่วงต้นจะมุ่งเน้นไปที่บทบาทของเซอร์ปินกับการเกิดโรคในมนุษย์ โดยเฉพาะภาวะพร่องแอลฟา-1 แอนตีทริปซิน (alpha1-antitrypsin (AAT) deficiency) ที่เป็นสาเหตุทำให้เกิดถุงลมโป่งพอง (emphysema) ถือเป็นหนึ่งในโรคทางพันธุกรรมที่พบได้บ่อยที่สุดในกลุ่มความผิดปกติที่เกี่ยวเนื่องกับเซอร์ปิน และภาวะพร่องแอนตีทรอมบิน (antithrombin deficiency) ที่ทำให้เกิดลิ่มเลือดอุดตัน ก็ได้รับความสนใจในการศึกษาเช่นเดียวกัน
ต่อมาในทตศวรรษที่ 1980 จากการศึกษาวิจัยทำให้ทราบได้แน่ชัดว่ากระบวนการยับยั้งเอนไซม์โปรตีเอสที่พบดังข้างต้นนั้น ส่วนหนึ่งเป็นผลมาจากโปรตีนที่เกี่ยวเนื่องกันในมหาวงศ์หนึ่ง ซึ่งมีทั้งชนิดที่ออกฤทธิ์เป็นสารยับยั้งเอนไซม์โปรตีเอส (เช่น แอลฟา-1 แอนตีทริปซิน) และชนิดที่ไม่มีฤทธิ์ยับยั้งเอนไซม์ (เช่น โอแวลบูมิน) จึงได้มีการตั้งชื่อกลุ่มโปรตีนนี้ว่า เซอร์ปิน โดยมีที่มาจากกลไกการออกฤทธิ์ที่พบได้บ่อยที่สุดของโปรตีนในมหาวงศ์นี้ (serine protease inhibitors). ในช่วงเวลาใกล้เคียงกัน ได้มีการวิเคราะห์โครงสร้างของเซอร์ปินชนิดแรกได้สำเร็จ (ครั้งแรกในรูปแบบอิสระ และครั้งที่ 2 ในรูปแบบที่เกิดการเปลี่ยนโครงรูปกับเอนไซม์) โดยโครงสร้างดังกล่าวแสดงให้เห็นว่า กลไกการออกฤทธิ์ยับยั้งการทำงานของเอนไซม์ของเซอร์ปินั้นมีส่วนเกี่ยวข้องกับการเปลี่ยนโครงรูปที่ผิดปกติของตำแหน่งกัมมันต์ การค้นพบนี้ส่งผลให้เกิดความสนใจในการศึกษาวิจัยเกี่ยวกับโครงสร้างของเซอร์ปินเพิ่มมากขึ้นในภายหลัง
ในปัจจุบัน มีการค้นพบเซอร์ปินแล้วมากกว่า 1,000 ชนิด ประกอบไปด้วย พบในมนุษย์ 36 ชนิด และที่เหลือพบในสิ่งมีชีวิตอื่นในอาณาจักรต่าง ๆ ทั้งสัตว์ พืช เห็ดรา แบคทีเรีย อาร์เคีย และไวรัสบางชนิด ในคริสต์ทศวรรษ 2000 มีการนำระบบการตั้งชื่อมาใช้เพื่อจัดหมวดหมู่โปรตีนชนิดต่าง ๆ ในมหาวงศ์เซอร์ปิน โดยอิงตามความสัมพันธ์ด้านวิวัฒนาการของเซอร์ปินแต่ละชนิด จึงถือได้ว่าเซอร์ปินเป็นกลุ่มโปรตีนที่ใหญ่และมีความหลากหลายที่สุดในบรรดาสารยับยั้งโปรตีเอส
วิวัฒนาการ
เซอร์ปินเป็นมหาวงศ์ของโปรตีนที่พบได้ทั่วไปและจัดเป็นกลุ่มโปรตีนยับยั้งโปรตีเอสที่ใหญ่ที่สุด แรกเริ่มเชื่อกันว่าสามารถพบเซอร์ปินได้ในสิ่งมีชีวิตประเภทยูแคริโอตเท่านั้น แต่หลังจากนั้นมาก็เริ่มมีการค้นพบเซอร์ปินในสิ่งมีชีวิตอื่นที่ไม่ใช่ยูแคริโอต อาทิ แบคทีเรีย อาร์เคีย และไวรัสบางชนิด แต่ก็ยังไม่สามารถทราบได้แน่ชัดว่า ยีนที่ทำหน้าที่สร้างเซอร์ปินที่พบในโพรแคริโอตนั้น เป็นยีนที่ได้รับการถ่ายทอดโดยรุ่นสู่รุ่นจากเซลล์โพรแคริโอตโบราณ หรือเป็นยีนที่ได้รับผ่านการถ่ายทอดยีนในแนวราบจากยูแคริโอต ทั้งนี้เซอร์ปินภายในเซลล์ส่วนใหญ่ (intracellular serpins) มีที่มาจากวงศ์วานวิวัฒนาการเคลดเดียวกัน ไม่ว่าจะเป็นชนิดที่พบในสัตว์หรือพืชก็ตาม จึงบ่งชี้ได้ว่า ทั้งเซอร์ปินภายในเซลล์และเซอร์ปีนภายนอกเซลล์ (extracellular serpins) นั้น อาจมีความหลากหลายมาก่อนที่จะเกิดการแยกเคลดวงศ์วานวิวัฒนาการของพืชและสัตว์ ยกเว้น ฮีตช็อกโปรตีน 47 ภายในเซลล์ (intercallular heat shock protein; HSP47) ซึ่งเป็นเซอร์ปินประเภทแชพเพอโรนที่มีความสำคัญต่อการม้วนพับตัวของคอลลาเจนและการเกิดห่วงโซ่ปฏิกิริยาระหว่างซิสเทอร์นี (cisternae) ของกอลไจแอปพาราตัสกับร่างแหเอนโดพลาซึม
การยับยั้งโปรตีเอสของโปรตีนเซอร์ปินนั้นถือได้ว่าเป็นกลไกที่มีมาตั้งแต่แรกเริ่ม ส่วนเซอร์ปินที่ไม่มีผลยับยั้งโปรตีเอสนั้นเป็นผลมาจากการวิวัฒนาการของโครงสร้างเพื่อพัฒนาหน้าที่ใหม่ (neofunctionalization) นอกจากนี้ เซอร์ปินที่ทำหน้าที่เป็นโปรตีนขนส่งบางชนิด อาจเกิดการเปลี่ยนโครงรูปจากไอโซเมอร์ชนิด S เป็นชนิด R เพื่อให้มีความจำเพาะในการเข้าจับกับตำแหน่งเป้าหมายมากยิ่งขึ้น
หน้าที่

โดยส่วนมากแล้ว โปรตีนในมหาวงศ์เซอร์ปินจะออกฤทธิ์ต้านซีรีนโปรตีเอสที่มีตำแหน่งออกฤทธิ์ภายนอกเซลล์และมีโครงสร้างคล้ายไคโมทริปซิน (chymotrypsin-like serine proteases) โดยโปรตีเอสเหล่านี้จะเข้าจับกับตำแหน่งเป้าหมายโดยใช้บซีรีนบริเวณ catalytic triad ของตำแหน่งกัมมันต์เข้าทำปฏิกิริยา ตัวอย่างโปรตีเอสเหล่านี้ได้แก่ ทรอมบิน ทริปซิน และ นิวโทรฟิลอีลาสเทส โดยเซอร์ปินจะออกฤทธิ์เป็นตัวยับยั้งสังหารแบบไม่สามารถผันกลับได้ (irreversible, suicide inhibitors) ต่อโปรตีเอสเหล่านั้น โดยการดักจับตัวกลางของกลไกการเร่งปฏิกิริยาของโปรตีเอส
อย่างไรก็ตาม ยังมีเซอร์ปินบางชนิดที่มีคุณสมบัติยับยั้งการทำงานของโปรตีเอสกลุ่มอื่น ๆ ส่วนใหญ่มักเป็นเอนไซม์ในกลุ่มซิสตีอีนโปรตีเอส จึงเรียกเซอร์ปินกลุ่มนี้ว่าเป็น "สารยับยั้งข้ามกลุ่ม" (cross-class inhibitors) โดยเอนไซม์ซิสตีอีนโปรตีเอสนั้นมีซิสตีอีนเป็นองค์ประกอบสำคัญในนิวคลีโอไฟล์ซึ่งต่างจากซีรีนโปรตีเอสที่มีนิวคลีโอไฟล์เป็นซีรีนเป็นตำแหน่งกัมมันต์ ถึงกระนั้น คุณสมบัติทางเคมีของเอนไซม์ทั้งสองก็ไม่ได้มีความแตกต่างกัน และกระบวนการการยับยั้งการทำงานของเอนไซม์โปรตีเอสทั้งสองกลุ่มโดยไซรีนนั้นก็ล้วนเกิดผ่านกลไกเดียวกัน ตัวอย่าง เซอร์ปินที่จัดเป็นสารยับยั้งข้ามกลุ่ม เช่น เซอร์ปิน บี4 ซึ่งเป็นแอนติเจนต่อมะเร็งผิวหนังสะแควมัสเซลล์คาร์ซิโนมา (squamous cell carcinoma antigen 1; SCCA-1) และ เซอร์ปินชนิด myeloid and erythroid nuclear termination stage-specific protein (MENT) ในสัตว์ปีก โดยเซอร์ปินทั้ง 2 ชนิดที่กล่าวถึงดังข้างต้นจะออกฤทธิ์ยับยั้งการทำงานของซิสตีอีนโปรตีเอสที่มีโครงสร้างคล้ายพาเพอิน (papain-like cysteine protease)
ฤทธิ์ทางชีววิทยา
ฤทธิ์ต้านโปรตีเอส
เซอร์ปินที่พบในมนุษย์ประมาณ 2 ใน 3 นั้นมีเป้าหมายการออกฤทธิ์บริเวณภายนอกเซลล์ ซึ่งมักจะเข้ายับยั้งการทำงานของโปรตีเอสในกระแสเลือดเพื่อควบคุมสมดุลการทำงานของเอนไซม์เหล่านั้น ตัวอย่างเช่น แอนติทรอมบินที่ออกต้านการทำงานของทรอมบินในกระบวนการการจับลิ่มของเลือด; แอนติทริปซิน, แอนตีไคโมทริปซิน และตัวยับยั้ง C1 ที่ออกฤทธิ์ในกระบวนการอักเสบและการตอบสนองของภูมิคุ้มกัน รวมไปถึง PAI-1 ในกระบวนการการซ่อมแซมเนื้อเยื่อที่เสียหาย นอกเหนือจากการยับยั้งวงจรการถ่ายโอนสัญญาณของโปรตีเอสแล้ว เซอร์ปินยังส่งผลต่อการพัฒนาของเนื่อเยื่อหรืออวัยวะเป้าหมายได้ด้วย ตารางแสดงเซอร์ปินที่พบในมนุษย์ (ด้านล่าง) แสดงให้เห็นถึงหน้าที่ของเซอร์ปินชนิดต่าง ๆ รวมไปถึงความผิดปกติที่เกิดขึ้นจากการขาดเซอร์ปินชนิดนั้น ๆ
ส่วนบทบาทของเซอร์ปินแต่ละชนิดที่ออกฤทธิ์ยับยั้งโปรตีเอสภายในเซลล์นั้นยังไม่สามารถระบุแยกได้แน่ชัด เนื่องจากมีการทำงานที่ซ้อนทับกันของเซอร์ปินหลายชนิดในกลไกหนึ่ง ๆ นอกจากนี้ เซอร์ปินหลายชนิดที่พบในมนุษย์นั้นยังไม่สามารถหาเซอร์ปินเทียบเคียงที่มีบทบาทหน้าที่สมมูลย์กันในสัตว์ทดลองได้ อย่างไรก็ตาม คาดว่าหน้าที่สำคัญของเซอร์ปินที่ออกฤทธิ์ภายในเซลล์นั้น อาจช่วยป้องกันการเกิดกิจกรรมที่ไม่เหมาะสมของโปรตีเอสภายในเซลล์ ตัวอย่างเช่น เซอร์ปินภายในเซลล์ของมนุษย์ที่เป็นที่คุ้นเคยกันมากที่สุดอย่าง เซอร์ปิน บี9 โดยเซอร์ปินชนิดนี้จะออกฤทธิ์ต้านการทำงานของแกรนูลที่บรรจุแกรนไซม์ บี ซึ่งเป็นโปรตีเอสที่เป็นพิษต่อเซลล์ ทำให้ช่วยป้องกันการปล่อยแกรนไซม์บีโดยไม่ได้ตั้งใจและลดการกระตุ้นการเกิดอะพอพโทซิสของเซลล์ก่อนเวลาอันควรหรือโดยไม่พึงประสงค์
นอกจากนี้ยังพบว่า ไวรัสบางชนิดมีการใช้เซอร์ปินเพื่อขัดขวางการทำงานของโปรตีเอสในโฮสต์ เช่น ไวรัสฝีดาษวัว (cowpox) ซึ่งจะสร้าง CrmA (cytokine response modifier A) ขึ้นมาเมื่อเข้าสู่เซลล์โฮสต์ ทั้งนี้เพื่อป้องกันไม่ให้เกิดการอักเสบและอะพอพโทซิสของเซลล์ที่ติดเชื้อ โดย CrmA จะทำให้เซลล์เป้าหมายมีการติดเชื้อเพิ่มมากขึ้นได้จากการกดกระบวนการอักเสบของเซลล์โฮสต์ด้วยการยับยั้งการทำงานของซิสตีอีนโปรตีเอสที่ชื่อแคสเปส 1 ส่งผลให้ไม่เกิดการเปลี่ยนโปรอินเตอร์ลิวคิน 1 และโปรอินเตอร์ลิวคิน 18 เป็นอินเตอร์ลิวคิน 1 (Interleukin 1; IL-1) และ อินเตอร์ลิวคิน 18 (Interleukin 18; IL-18) ตามลำดับ ซึ่งทั้งสารทั้ง 2 ชนิดนี้จัดเป็นไซโตไคน์ที่จำเป็นต่อการเกิดไพรอพโทซิสของเซลล์ ส่วนในยูแคริโอตนั้น พบว่าเซอร์ปินของพืชสามารถยับยั้งได้ทั้งกลุ่มเอนไซม์เมตาแคสเปส และซิสตีอีนโปรตีเอสที่มีโครงสร้างคล้ายพาเพอิน
บทบาทอื่น
เซอร์ปีนภายนอกเซลล์ที่ไม่มีผลยับยั้งโปรตีเอสนั้นจะทำหน้าที่อื่นที่หลากหลายแตกต่างอันออกไป อาทิ ไทโรซีน-บายดิ้ง กลอบูลิน และทรานส์คอร์ติน ซึ่งทำหน้าที่ขนส่งฮอร์โมนไทรอกซีน และคอร์ติซอล ตามลำดับ; โอแวลบูมิน ซึ่งเป็นโปรตีนที่พบมากที่สุดในไข่ขาว ก็จัดเป็นเซอร์ปีนอีกชนิดหนึ่ง ถึงแม้จะไม่ทราบบทบาทหน้าที่ที่แน่ชัดเท่าใดนัก แต่คาดว่าโอแวลบูมินนี้อาจทำหน้าที่เป็นโปรตีนเก็บสะสมสำหรับการพัฒนาการก่อนการเกิดของตัวอ่อน (Prenatal development) รวมไปถึงฮีตช็อกโปรตีน 47 ซึ่งจัดเป็นเซอร์ปินประเภทแชพเพอโรนที่มีความสำคัญต่อการม้วนพับตัวของคอลลาเจน และช่วยให้เกลียวสามสายของคอลลาเจนมีความเสถียรระหว่างกระบวนการสังเคราะห์คอลลาเจนในร่างแหเอนโดพลาซึม
อย่างไรก็ตาม ยังมีเซอร์ปินบางชนิดที่นอกจากจะออกฤทธิ์ต้านโปรตีเอสแล้ว ยังมีหน้าที่อื่นที่ไม่เป็นผลเกี่ยวเนื่องมาจากการยับยั้งโปรตีเอส ตัวอย่างเช่น MENT ซึ่งเป็นเซอร์ปินที่พบในนก นอกจากจะออกฤทธิ์ต้านซิสตีอีนโปรตีเอสภายในเซลล์แล้ว ยังเป็นสารที่มีส่วนช่วยในการดัดแปรโครมาติน (chromatin remodeling) ภายในเซลล์เม็ดเลือดแดงของนกอีกด้วย
โครงสร้าง
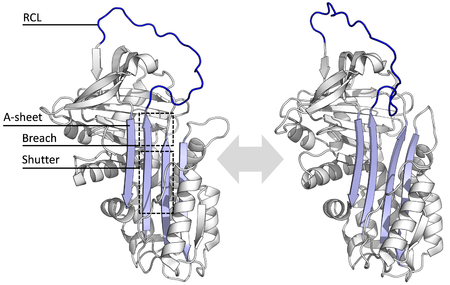
เซอร์ปินทุกชนิดจะมีโครงสร้างพื้นฐานที่เหมือนกัน ถึงแม้จะมีหน้าที่ที่หลากหลายแตกต่างกันก็ตาม โครงสร้างดังกล่าวจะประกอบไปด้วยแผ่นบีตา 3 แผ่น (ชื่อว่าแผ่น A, B และ C) และเกลียวแอลฟาจำนวน 8–9 เกลียว (ชื่อว่าเกลียว hA–hI) โดยส่วนที่มีความสำคัญอย่างมีนัยยะต่อการทำงานของเซอร์ปินได้แก่ แผ่น A-sheet และวงศูนย์ปฏิกิริยา (reactive centre loop; RCL) ซึ่งแผ่น A-sheet นี้จะประกอบไปด้วยสายบีตาจำนวน 2 สายเรียงตัวในแนวขนานกัน พื้นที่ที่อยู่ระหว่างสายบีตาทั้ง 2 สายนี้ เรียกว่า 'ซัตเตอร์' (shutter) และส่วนที่ด้านบนขึ้นไปเรียกว่า 'บรีช' (breach) โดยโครงสร้างที่เป็นวงศูนย์ปฏิกิริยาจะเป็นส่วนที่เริ่มเข้าจับกับโปรตีเอสเป้าหมายเพื่อยับยั้งการทำงานของโปรตีเอสนั้น ๆ จากโครงสร้างของเซอร์ปินที่ได้จากการจำลองพบว่าวงศูนย์ปฏิกิริยาของเซอร์ปีนจะอยู่ทั้งในระยะเครียดเต็มที่ (fully stressed) หรือในระยะคลายบางส่วน (partially relaxed) ซึ่งบางส่วนของวงจะม้วนพับเข้าไปในเกลียว A-sheet คาดการณ์กันว่าสมดุลไดนามิก (dynamic equilibrium) ของเซอร์ปินนั้นจะอยู่ระหว่าง 2 ระยะนี้ นอกจากนี้วงศูนย์ปฏิกิริยาดังกล่าวยังสามารถเกิดปฏิกิริยาแบบชั่วคราวกับโครงสร้างส่วนที่เหลือของเซอร์ปินได้ เนื่องจากโครงสร้างส่วนนี้มีความยืดหยุ่นสูงและสัมผัสอยู่กับสารละลายอยู่ตลอดเวลา
ในปัจจุบันมีการค้นพบโครงสร้างของเซอร์ปินครอบคลุมหลากหลายโครงรูป ซึ่งถือเป็นส่วนสำคัญในการศึกษาเพื่อทำความเข้าในในกลไกการออกฤทธิ์ที่สลับซับซ้อนของโปรตีนกลุ่มนี้ ดังนั้น การศึกษาชีววิทยาโครงสร้างจึงได้เข้ามามีบทบาทสำคัญในการทำความเข้าใจเกี่ยวกับการทำงานและฤทธิ์ทางชีววิทยาของเซอร์ปิน
การกระจาย
สัตว์
มนุษย์
จีโนมของมนุษย์ที่สามารถถอดรหัสเพื่อสร้างเซอร์ปินได้นั้นมีทั้งหมด 16 เคลด กำหนดให้มีชื่อตั้งแต่ serpinA ถึง serpinP โดยทั้งหมดนี้สามารถถอดรหัสเพื่อสร้างเซอร์ปินชนิดยับยั้งและชนิดไม่ยับยั้งได้ 29 และ 7 ชนิด ตามลำดับ ระบบการตั้งชื่อเซอร์พินของมนุษย์นั้นขึ้นอยู่กับการวิเคราะห์ทางวิวัฒนาการของเซอร์ปิน ซึ่งตั้งแต่ปี ค.ศ. 2001 เป็นต้นมา มีการค้นพบเซอร์ปินในมนุษย์แล้วประมาณ 500 ชนิด โดยการกำหนดชื่อจะใช้รหัส serpinXY โดยที่ X คือ เคลดของโปรตีนและ Y จำนวนของโปรตีนภายในเคลดนั้น ซึ่งบทบาทหน้าที่ของเซอร์ปินแต่ละชนิดที่ค้นพบในมนุษย์ได้วิเคราะห์ได้จากการศึกษาทางชีวเคมี, ความผิดปกติทางพันธุกรรมของมนุษย์ และจากการศึกษาในหนูน็อกเอาต์ (knockout mouse)
| ตารางแสดงเซอร์ปินที่พบในมนุษย์ | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
|
เซอร์ปินพิเศษของสัตว์เลี้ยงลูกด้วยนม
เซอร์ปินหลายชนิดที่พบในสัตว์เลี้ยงลูกด้วยนมพบว่าไม่สามารถเทียบเคียงหาเซอร์ปีนที่มีหน้าที่และฤทธิ์ทางชีววิทยาที่ทัดเทียมกันได้ ตัวอย่างเช่นเซอร์ปินที่พบในสัตว์ฟันแทะหลายชนิด (โดยเฉพาะอย่างยิ่งเซอร์ปินชนิดออกฤทธิ์ภายในเซลล์ของสัตว์ในวงศ์ย่อยหนู) รวมไปถึงยูเทอรีนเซอร์ปินซึ่งจัดหนึ่งในสมาชิกของเคลด A (SERPINA) โดยเซอร์ปินชนิดนี้จะถูกถอดรหัสยีน SERPINA14 และถูกสร้างขึ้นโดยเซลล์เยื่อบุโพรงมดลูกของสัตว์เลี้ยงลูกด้วยนมอันดับสัตว์กีบคู่ภายใต้อิทธิพลของโพรเจสเทอโรนและเอสโตรเจน ซึ่งเซอร์ปินเหล่านี้อาจไม่ได้มีหน้าที่ในการต้านโปรตีเอส และอาจจะมีบทบาทในช่วงการตั้งครรภ์ ทั้งนี้เพื่อยับยั้งการตอบสนองของระบบภูมิคุ้มกันมารดาไม่ให้ต่อต้านเนื้อเยื่อต่าง ๆ ที่ถูกสร้างขึ้นระหว่างการตั้งครรภ์ (conceptus) หรืออาจมีส่วนในการช่วยขนส่งสารต่าง ๆ ทางรก
แมลง
ในจีโนมของแมลงวันทองมียีนมากถึง 29 โลคัสที่ทำหน้าที่ในการสร้างเซอร์ปิน ผลการวิเคราะห์ลำดับกรดอะมิโนพบว่า เซอร์ปิน 14 ชนิดถูกควบคุมโดยเคลด Q และ 3 ชนิดโดยเคลด K ส่วนเซอร์ปินที่เหลืออีก 12 ชนิด จัดเป็นเซอร์ปินกำพร้าที่ไม่จำเพาะต่อเคลดใด ๆ ระบบการจำแนกประเภทเคลดที่ใช้กับมนุษย์นั้นไม่สามารถใช้กับกรณีนี้ได้ดีเท่าใดนัก จึงได้นำเอาระบบการตั้งชื่อมาประยุกต์ใช้แทน โดยอิงตามตำแหน่งของยีนบนโครโมโซมที่ควบคุมการสร้างเซอร์ปินนั้น ๆ แทน ซึ่งพบว่าเซอร์ปิน 13 ชนิดที่แยกได้นั้นถูกควบคุมโดยยีนที่อยู่คนละตำแหน่งกัน (รวมถึง Serpin-27A ที่แสดงรายละเอียดด้านล่าง) และที่เหลืออีก 16 ชนิดถูกควบคุมโดยกลุ่มยีน 5 ยีนที่ตำแหน่ง 28D (2 เซอร์ปิน), 42D (5 เซอร์ปิน), 43A (4 เซอร์ปิน), 77B (3 เซอร์ปิน) และ 88E (2 เซอร์ปิน)
การศึกษาเกี่ยวกับเซอร์ปินของแมลงวันทองพบว่า Serpin-27A นั้นออกฤทธิ์ต้านอีสเตอร์โปรตีเอส (โปรตีเอสลำดับสุดท้ายในวัฎจักร Nudel, Gastrulation Defective, Snake and Easter) ที่ควบคุมการสร้างเนื้อเยื่อในอวัยวะส่วนแกนหลังและหน้าท้อง อีกทั้งยังมีส่วนช่วยในการทำงาน Spätzle (ลิแกนด์ประเภทไคโตไซน์ชนิดหนึ่ง) เพื่อกระตุ้นตัวรับ Toll-like receptor ในระบบภูมิคุ้มกันเพื่อให้ทนต่อแบคทีเรีย รา และไวรัส นอกจากนี้ การส่งสัญญาณ toll ที่เป็นผลจากการกระตุ้นตัวรับ Toll-like receptor ยังมีบทบาทสำคัญในการพัฒนาระบบภูมิคุ้มกันโดยกำเนิดของแมลงต่าง ๆ อีกด้วย
ในกระดิ่งเงินกระดิ่งทองมีโปรตีน SPN93 ซึ่งประกอบไปด้วยเซอร์ปิน 2 หน่วยย่อย ทำหน้าที่ควบคุมวัฎจักรการสลายโปรตีนผ่านการถ่ายทอดสัญญาณทอล (Toll signaling cascade)
หนอนตัวกลม
จีโนมของ C. elegans ซึ่งเป็นสัตว์จำพวกหนอนตัวกลม สามารถถอดรหัสสร้างเซอร์ปินได้ 9 ชนิด โดยเซอร์ปินทั้งหมดนี้ไม่มีเพปไทด์ส่งสัญญาณและดูคล้ายจะเป็นเซอร์ปินประเภทภายในเซลล์ อย่างไรก็ตาม มีเพียงเซอร์ปินเพียง 5 ชนิดเท่านั้นที่ดูเหมือนว่าจะสามารถออกฤทธิ์ยับยั้งการทำงานโปรตีเอสได้ หนึ่งในนั้นก็คือ SRP-6 ที่ทำหน้าที่ป้องกันและต้านทานการแตกของไลโซโซมที่สัมพันธ์กับคาลเพนและมีภาวะเครียดเป็นปัจจัยเหนี่ยวนำ (stress-induced calpain-associated lysosomal disruption) นอกจากนี้ SRP-6 ยังช่วยยับยั้งการทำงานของซิสตีอีนโปรตีเอสที่ถูกปล่อยออกมาหลังการแตกของไลโซโซม ด้วยเหตุนี้จึงทำให้ C. elegans ที่ขาด SRP-6 นั้น มีความไวต่อภาวะเครียดมากกว่าปกติ และพบว่าหนอนที่ถูกหยุดการทำงานของยีนที่ควบคุมการสร้าง SRP-6 (knockout worms) จะตายเกือบทันทีเมื่อถูกนำไปวางในน้ำ (ผลจาก hypo-osmotic stress lethal phenotype; Osl) ส่วนสาเหตุอื่น ๆ ที่ทำให้การแสดงออกของ SRP-6 ลดน้อยลงจนนำไปสู่การตายของเซลล์ เนื้อเยื่อหรือทั้งระบบ ได้แก่ ภาวะฉุกเฉินจากความร้อน ความเครียดออกซิเดชัน ภาวะพร่องออกซิเจน และ MET-4 จึงสรุปได้ว่า SRP-6 และไลโซโซมมีบทบาทสำคัญเป็นอย่างมากในการตอบสนองภาวะเครียดและการควบคุมตายของเซลล์ในหนอนตัวกลม
พืช
เซอร์ปินที่ถูกค้นพบเป็นกลุ่มแรก ๆ นั้นเป็นเซอร์ปินที่พบในพืช โดยเซอร์ปิน Z ที่มีอยู่มากในเมล็ดข้าวบาร์เลย์และเป็นหนึ่งในส่วนประกอบโปรตีนที่สำคัญที่พบในเบียร์ รวมไปถึงจีโนมของพืชต้นแบบอย่างเธลเครสที่มียีนที่คล้ายเซอร์ปินอยู่ 18 ยีน ถึงแม้ว่าจะมีเพียง 8 ยีนเท่านั้นที่มีลำดับนิวคลีโอไทด์ของเซอร์ปินยาวเต็มที่
การทดสอบในหลอดทดลอง (in vitro) พบว่า เซอร์ปินจากพืชบางชนิดออกฤทธิ์ยับยั้งซีรีนโปรตีเอสที่มีโครงสร้างคล้ายไคโมทริปซิน (chymotrypsin-like serine proteases) ของสัตว์เลี้ยงลูกด้วยนมได้เป็นอย่างดี ตัวอย่างการศึกษาที่อธิบายปรากฏการณ์ดังกล่าวได้ดีที่สุดนั้นเป็นการศึกษาการทำงานของเซอร์ปิน Zx (BSZx) ในข้าวบาร์เลย์ซึ่งพบว่ามีความสามารถในการต้านทริปซินและไคโมทริปซิน รวมไปถึงแฟกเตอร์ต่าง ๆ ที่เกี่ยวข้องกับการจับลิ่มของเลือดในมนุษย์หลายชนิด อย่างไรก็ดี เอนไซม์ที่มีคุณสมบัติใกล้เคียงซีรีนโปรตีเอสที่มีโครงสร้างคล้ายไคโมทริปซินนั้นไม่ปรากฏว่ามีอยู่ในพืช นอกจากนี้ยังพบว่าวงศูนย์ปฏิกิริยาของเซอร์ปินหลายชนิดที่พบในข้าวสาลีและข้าวไรย์มีการเรียงลำดับ PolyQ ต่อเนื่องซ้ำ ๆ คล้ายกับที่พบในโพรลามีนซึ่งเป็นโปรตีนเก็บสะสมภายในเอนโดสเปิร์มของเมล็ดพืช จึงอาจพอสรุปได้ว่า เซอร์ปินที่พบในพืชเหล่านี้อาจทำหน้าที่ในการยับยั้งโปรตีเอสจากแมลงหรือจุลชีพที่มีคุณสมบัติในการย่อยโปรตีนเก็บสะสมเมล็ด โดยมีการศึกษาหนึ่งที่ให้ข้อสรุปสนับสนุนสมมติฐานข้างต้น กล่าวคือ ถึงแม้จะพบความสัมพันธ์แบบแปรผกผันระหว่างการแสดงออกของ CmPS-1 ซึ่งเป็นเซอร์ปินที่พบในสารละลายสารอาหาร (phloem sap) ของต้นฟักทองและแตงกวา กับการอยู่รอดและการขยายพันธุ์ของเพลี้ยอ่อน แต่การศึกษาในหลอดทดลองกลับพบว่า การให้สารละลาย CmPS-1 ความบริสุทธิ์สูงที่สกัดจากสารละลายสารอาหารข้างต้น กลับไม่มีผลต่อการรอดชีวิตของกลุ่มตัวอย่าง
นอกจากนี้ ยังมีการค้นพบหน้าที่อื่นหรือโปรตีเอสเป้าหมายอื่นของเซอร์ปินพืชอีกหลายชนิด อาทิ เซอร์ปิน AtSerpin1 (At1g47710; 3LE2) ที่พบในพืชสกุลอะราบิดอบซิส ที่ทำหน้าที่เป็นสารตัวกลางในการควบคุมการเกิดไม่ให้เกิดการทำงานของโปรแกรมการตายของเซลล์ที่มากเกินไป โดยมีเป้าหมายการออกฤทธิ์ที่ซิสเตอีนโปรตีเอสชื่อว่า 'Responsive to Desiccation-21' (RD21) และการศึกษาในหลอดทดลองยังพบว่า AtSerpin1 สามารถยับยั้งเอนไซม์โปรตีเอสที่คล้ายเมตาแคสเปสได้อีกด้วย ทั้งนี้ ยังมีเซอร์ปินอีก 2 ชนิดที่พบในพืชดังกล่าว ได้แก่ AtSRP2 (At2g14540) และ AtSRP3 (At1g64030) ซึ่งจะทำงานตอบสนองต่อการได้รับความเสียหายของดีเอ็นเอ
เห็ดรา
ณ ปัจจุบัน มีการค้นพบเซอร์ปินในสิ่งมีชีวิตจำพวกเห็ดราเพียง 1 ชนิด คือ เซลปิน (celpin) จากรา Piromyces spp. สายพันธุ์ E2 ซึ่งเป็นราที่ไม่ต้องการออกซิเจน (anaerobic fungi) ในจีนัส Piromyces อาศัยอยู่ในทางเดินอาหารของสัตว์เคี้ยวเอื้อง โดยราชนิดนี้มีส่วนช่วยสำคัญในการย่อยสลายชิ้นส่วนของพืชที่สัตว์เหล่านี้กินเข้าไป ทั้งนี้ ปลายซี (C-terminus) ของเซลปินประกอบไปด้วยกรดอะมิโน 380 หน่วย ซึ่งเป็นส่วนที่แสดงคุณสมบัติเป็นเซอร์ปิน ส่วนปลายเอ็น (N-terminus) จะประกอบไปด้วยดอคเกอรินจำนวน 2 โมเลกุล ซึ่งเป็นสารที่มีส่วนช่วยในการสร้างเซลลูโลโซมของรา เพื่อใช้เป็นแหล่งสะสมเอนไซม์ที่จำเป็นในการย่อยชิ้นส่วนพืช จึงมีความเป็นไปได้ว่า เซลปินที่พบในราสายพันธุ์นี้อาจมีหน้าที่ป้องกันไม่ให้เซลลูโลโซมถูกทำลายด้วยโปรตีเอสจากพืช นอกจากนี้ยังมีรายงานการค้นพบเซอร์ปินของแบคทีเรียที่ทำหน้าที่คล้ายคลึงกันกับเซลปินอีกด้วย
โพรคาริโอต
ไวรัส
การเปลี่ยนโครงรูปและกลไกการยับยั้ง
การเสื่อมสลาย
โรคจากความผิดปกของเซอร์ปิน
ดูเพิ่ม
แหล่งข้อมูลอื่น
- PDB Molecule of the Month Serpin
- Merops protease inhibitor claudication (Family I4)
- Serpins ในหอสมุดแพทยศาสตร์แห่งชาติอเมริกัน สำหรับหัวข้อเนื้อหาทางการแพทย์ (MeSH)
- James Whisstock laboratory ของมหาวิทยาลัยโมนาช
- Jim Huntington laboratory เก็บถาวร 2016-10-30 ที่ เวย์แบ็กแมชชีน ของมหาวิทยาลัยเคมบริดจ์
- Frank Church laboratory ของมหาวิทยาลัยนอร์ทแคโรไลนา แชเปิลฮิลล์
- Paul Declerck laboratory ของ มหาวิทยาลัยคาทอลิคเลอเฟิน
- Tom Roberts laboratory ของมหาวิทยาลัยซิดนีย์
- Robert Fluhr laboratory ของ สถาบันวิทยศาสตร์ไวซ์มานน์
- Peter Gettins laboratory ของมหาวิทยาลัยอิลลินอยส์ ชิคาโก
- ข้อมูลเชิงโครงสร้างทั้งหมดของ UniProt: P01009 (Human Alpha-1-antitrypsin) สามารถเข้าถึงได้ในธนาคารข้อมูลโปรตีนของ PDBe-KB.